
GRAFTING IN ANIMALS. Every gardener is well ac quainted with grafting in plants. But it is less well-known that pieces of animals too may be joined in permanent union. Grafting in animals is practised mainly for scientific purposes or for the restoration of weakened or lost parts. Therefore graft and stock are not always taken from different species (heteroplastic trans plantation) or races (alleloplastic), but may belong to samples of the same species and race (homoplastic) or even to one individual (autoplastic). It is as a rule easier to join pieces of the same species than pieces belonging to different species. In the warm blooded animals difficulties may arise even when two individuals of the same species are united by grafting, owing to blood incom patibility. With adult specimens of cold-blooded vertebrates this difficulty is less, and in their earlier stages and in invertebrate animals there seems to be little inconvenience from this source. In some types it is possible to join species belonging to different classes (dysplastic transplantation), e.g., amphibians and fish. The degree to which grafting may be carried on depends, too, on the injury a given animal can endure. Developed vertebrates will not stand the stoppage of their circulation and breathing in evitable with the removal of the head. But in embryos of frogs, before the circulation of blood has started, the head-region may be grafted on to the body even of another species, and such com pound monsters may even pass metamorphosis. As insects do not need their heads for breathing, and circulation continues without the head being present, grafting of the head is possible even in the imaginal instar, but it is not yet clear whether function is restored. There is no doubt that in lower organisms, such as worms, the head or other body-region can be grafted with functional success. Lengthened individuals may be obtained in embryos of amphib ians, tadpoles of abnormal length resulting. Two warm-blooded animals may be joined by bridges of tissue containing blood vessels and nerves. Side-to-side "Parabiosis" of this kind has been effected in rats, and reminds one of the "Siamese twins." Para biotic grafts have also been effected in frogs, newts, butterfly pupae, earthworms and hydrae. .

In certain cases the technique of grafting is even simpler in animals than in plants ; when the opportunity is given for the stock to grip the graft by its own means, not even tying or gluing is required (autophoric method). Thus the sea-lily Antedon, a relative of the starfish, bears a central disc easily detached. If another disc taken, let us say, from a specimen of different colour, be inserted into the groove left after the removal of the central disc, it will be clutched and kept in place by the tentacles sur rounding this spot. There is nothing mysterious about this re action as the tentacles also fold over the disc in the normal animal. In the case of the vertebrate eye an implanted eye-ball will keep in place by friction and the closing of the eyelids. To prevent these from opening prematurely, a fine pin or a silk stitch can be applied. Eyes can also be grafted in forms (e.g., fish and tad poles), which have no eyelids and have no means of pawing at the replanted eyes. Replantation of eyes has been successfully achieved in several species of fresh-water fish, newts, salamanders, frogs, toads, rats and in a rabbit. But only in very rare cases does sight return. Endeavours to apply grafting of eyes to restore vision to domestic animals have hitherto been unsuccessful. The lens alone may be grafted autophorically. The lens is extracted as in the operation for cataract. In fish, with cataract, the damaged lens, easily recognized by its opacity, is removed and a transparent lens from the eye of a healthy fish is slipped through the slit made by the operator's knife and is retained by the cornea. Transplantation of the lens in warm-blooded animals has not yet been recorded. Grafting of limbs, arm or leg, may be done in young tailed amphibians by inserting freshly detached limbs between the muscles of the shoulder pit or inguinal region, the contraction of the muscles holding them in place. In warm blooded animals, however, this method is not practicable and one has to resort to uniting every blood-vessel and nerve-trunk by stitching with catgut or silk. A special technique has been devised for this purpose. The suturing of nerves, however, in trans planting mammals' limbs has not proved satisfactory; no return of motility being secured. In cold-blooded animals sewing is widely used. Chrysalids of butterflies and moths have been united by girdles of paraffin after cutting on ice. Small sea or fresh water animals can be fixed in grooves of wax and covered by slips of glass or metal (silver, lead) so that the two cut ends touch each other; they then become united by the pressure. In this manner tadpoles, planarians and hydras may be dealt with. A convenient method of joining two or more pieces of hydra has been found by threading them on fine bristle. When the thread is taken out of the water, a drop of water remains on the grafts and draws them together by surface-tension.
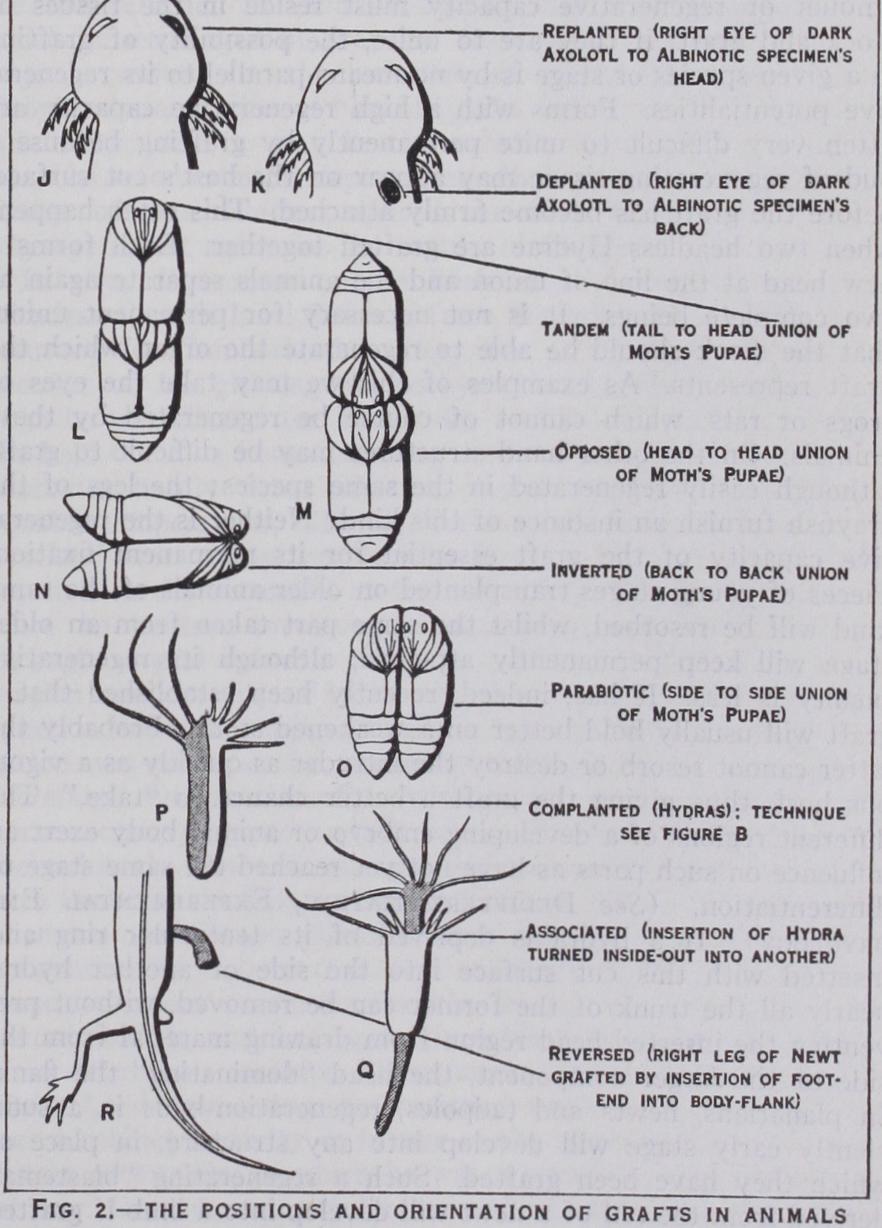
Antagonism of Graft and Re-growth.—Although a certain amount of regenerative capacity must reside in the tissues of stock and graft, if they are to unite, the possibility of grafting in a given species or stage is by no means parallel to its regenera tive potentialities. Forms with a high regenerative capacity are often very difficult to unite permanently by grafting because a bud of regenerating tissue may appear on the host's cut surface, before the graft has become firmly attached. This often happens when two headless Hydrae are grafted together. Each forms a new head at the line of union and the animals separate again as two complete beings. It is not necessary for permanent union, that the stock should be able to regenerate the organ which the graft represents. As examples of this we may take the eyes of frogs or rats, which cannot of course be regenerated by these animals. On the other hand structures may be difficult to graft, although easily regenerated in the same species : the legs of the crayfish furnish an instance of this kind. Neither is the regenera tive capacity of the graft essential for its permanent fixation. Pieces of young stages transplanted on older animals of the same kind will be resorbed, whilst the same part taken from an older stage will keep permanently attached, although its regenerative faculty is less. It has, indeed, recently been established that a graft will usually hold better on a weakened stock. Probably the latter cannot resorb or destroy the intruder as quickly as a vigor ous host, thus giving the graft a better chance to "take." The different regions of a developing embryo or animal body exert an influence on such parts as have not yet reached the same stage of differentiation. (See DEDIFFERENTIATION, EXPERIMENTAL EM