
FUNGI, a large group of plants devoid of green colouring matter (chlorophyll) and reproduced by spores. They include many of the lowest forms of plant life. The thallus is unicellular or composed of branched tubes or filaments which have apical growth. The spore may consist of one or many cells. The absence of chlorophyll has a striking effect on the mode of life of fungi since in its absence they are unable to synthesise carbohydrates from the carbon dioxide of the air. They are dependent therefore on other plants or animals for their supplies of carbohydrates and some times of organic compounds of nitrogen. These supplies are ob tained from living organisms by parasitic fungi, or more com monly from their dead remains by fungi which are termed sapro phytes. Certain non-chlorophyllous organisms however are sepa rated from the fungi and regarded as independent groups.
The fungi are a large group distributed over the whole world wherever other living organisms are found on which they may depend for their subsistence. The number of species is very large; probably 1o0,000 is a conservative estimate, but no figures, however carefully compiled, can be other than provisional. The bulk of the species known belong to the temperate zone. A wealth of forms no doubt exists in the tropics which have yet to be dis covered. Geologically the fungi are an old group; the delicate nature of their tissues has militated against their preservation as fossils, but the hyphae of parasitic fungi are found in some of the earliest plant fossils known to us, the beautifully preserved re mains of the Rhynie chert of Devonian age.

Organization.—If spores of common white mould (Mucor) are placed in a suitable food-solution they readily germinate ; they absorb water and swell—the dark outer coat (exospore) rup tures and the thin colourless inner coat (endospore) protrudes and lengthens to form a short tube, known as the germ tube (fig. I c). This tube grows at its tip and lengthens considerably; branches arise from it, elongate and branch in their turn and so give rise to an interconnected series of tubular structures known as hyphae. Each hypha is covered externally by a membrane and encloses a mass of protoplasm in which many nuclei are embedded. In older hyphae vacuoles appear and coalesce to form a large cen tral vacuole. Through the walls of the hyphae food solutions are absorbed which provide for their further growth and extension. The whole system of hyphae thus represents the vegetative body of the fungus known as the mycelium (fig. i a). As the develop ment of the mycelium proceeds the work of absorption becomes restricted to the younger hyphae which are growing forward into fresh areas of the substratum. The protoplasm of the older parts of the mycelium becomes stored with reserve food-materials and sooner or later from this part of the mycelium arises a new kind of hypha, devoted to the reproduction of the mould. These hyphae are stouter, limited in growth and with a different reaction to such stimuli as light, gravity, oxygen, etc. Thus they normally grow upwards into the air, towards the light, and terminate their growth by swelling at the tip to form a spherical head (the sporan gium), the contents of which divide into many portions, each of which secretes a wall and becomes a spore (fig. 1 b) . The produc tion of sporangia continues while external circumstances remain favourable and while fresh supplies of food material are available to the developing mycelium. Given these conditions there is no reason why growth should ever cease. In the case of the fungi causing the well-known fairy rings, common in pastures, from a comparison of the rate of growth with the size of the ring, it has been estimated that the age of certain rings is not less than 3-400 years. Although the actively growing hyphae of Mucor are con tinuous tubes with numerous nuclei in their protoplasm and are hence described as caenocytic, the older hyphae show numerous cross walls which serve to cut off useless side-branches and to concentrate the food-reserves. In the majority of fungi all the hyphae are segmented (multicellular), the segments being mul tinucleate or more commonly with one or two nuclei.
The cell walls of certain groups of fungi (Saprolegniaceae and Peronosporaceae) give the same reactions as those of the higher plants and apparently consist of cellulose, but Gilson and Van Wisselingh have shown that in a very large number of forms the main constituent of the wall is chitin, a substance common in many animals, e.g. insects. In many cases the membranes undergo secondary changes and other substances may be added such as resins and colouring matters, the walls may become thickened by the deposition of new layers and often profoundly altered by ligni fication and suberization (Fomes, Daedalea). A striking change is carbonization, in which the walls become black and brittle.
(Daldinia, Cucurbitaria).—In the reproductive bodies of many of the higher fungi the walls of certain hyphae become mucilagi nous, being hard and horny when dry but capable of absorbing water with great rapidity and consequent swelling. Striking thickening and differentiation may occur in the walls of spores which are commonly pitted, sculptured and ridged.
In the simpler types of mycelium the hyphae show little differ entiation except in size. In many parasites in which the hyphae traverse the intercellular spaces of their hosts, lateral branches are produced which penetrate into the living cells and there carry on the absorption of food materials, these haustoria may have the form of simple or branched hyphae or swollen vesicles. In many mycelia the hyphae may become woven together into threads or cords. These may be white and tender as in the "spawn" of the common mushroom, or develop an outer hard and dark coloured layer and so assume a root-like appearance as in the rhizomorphs of the honey fungus, Armillaria mellea. In other cases the closely interwoven hyphae may form compact masses, varying in size from that of a pin's head (Botrytis) to that of a man's head and weigh ing 10 to 25 lb. (Polyporus Myllitae). These bodies known as sclerotia show also a differentiation into a hard, black outer layer and an inner white core—the whole body consisting of hyphae so closely interwoven and copiously branched and segmented as to resemble in section parenchymatous tissue of one of the higher plants. From its mode of formation, however, this tissue is more properly described as plectenchyma or pseudo-parenchyma.
Both rhizomorphs and sclerotia serve for the collection and storage of reserve foods to be used in the formation of reproduc tive bodies, or in the resumption of vegetative activity. The former also serve to extend the fungus and in some parasites act as organs of attack (Armillaria).
In the higher fungi a similar interweaving and compacting of hyphae are involved in the formation of the fruit bodies or sporo phores and in these structures a high degree of elaboration and of differentiation of the plectenchyma may occur (Polyporaceae, Phallaceae fig. 19).
The particular mode of nutrition of the fungi, viz., their de pendence on other organisms for their food materials, is only rendered possible by their power of secreting a whole range of ferments capable of digesting the many complex substances which go to form the bodies of plants and animals. Amongst the enzymes extracted from fungi may be mentioned as examples, diastase which converts starches into sugar, cytase with a similar action on cellulose, peptase which converts proteins into peptones and amides, lipase which hydrolyses fat and oils, invertase which splits cane sugar into simple sugars such as grape sugar, and zymase which decomposes grape sugar into alcohol and carbon dioxide. These or similar enzymes are found as normal products of metabolism in plants generally where they play their part in the necessary digestion of reserve foods, etc. Whilst, however, in the higher plants their work is performed with few exceptions in the cells or the tissues in which they arise, in the fungi these ferments are excreted by the hyphae in order to bring about the necessary changes in the food-substance with which they come into contact. All fungi are apparently not capable of secreting every kind of ferment and so are unable to utilize all classes of food materials. Many are restricted in their choice, or rather show special facility in dealing with particular materials, e.g., horn, feathers, cellulose, fats. Some moulds on the other hand (Penicilliurn, Aspergillus) can utilize almost any organic food material and a large number of enzymes have in fact been extracted from these forms.
Although the small nuclei of certain fungi were not recognized by early investigators, subsequent improvements in methods of research have demonstrated their presence in all the species in which they have been sought. In the lower fungi the hyphae are coenocytic, i.e., they contain numerous nuclei and these are of small size (.001-3 m.m. in the Mucoraceae). In the higher fungi the segments may contain several nuclei (Pyronema, Aspergillus) but usually have only one or two. There is no reason to doubt that not only are they as essential to the cell of the fungus as to that of any other organism, but that they are fully comparable with those of the higher plants both in essential structure and in function. They normally divide mitotically (see CYTOLOGY) and in a number of cases, where the nuclei are of considerable size, the details of division have been minutely described (Phyllactinia, Humaria). Typical spindles with centrosomes and asters are formed and the chromosomes are clearly differentiated. The num ber of chromosomes is usually small, e.g., eight in Phyllactinia.
As in the higher plants, in those fungi in which sexual repro duction occurs, the essential feature of fertilization is the union of two nuclei. These are, in the simplest cases, the nuclei of the sexual cells or gametes and the new cell which results is known as the zygote. This has a double number of chromosomes, and is therefore described as diploid in contradistinction to the haploid nuclei of the sexual cells and of any vegetative cells associated with them. Fertilization thus marks the end of one definite phase in the life of the fungus (haploid phase) and the beginning of a new one (diploid phase) of which the zygote is the starting point. This in its turn must end with a process of reduction (meiosis) by which the haploid nuclei are again restored (see CYTOLOGY). Fertilization and meiosis are thus two cardinal points in the history of an organism ; ending the haploid and diploid phases respectively. In some of the higher fungi, these alternating nuclear phases are associated and correlated with striking differ ences in the vegetative structure of the mycelia concerned. In these the zygote may give rise to many generations of diploid cells which eventually produce the mother-cells—usually of special form—in which meiosis will occur. These cells—gonoto conts—correspond to the spore-mother-cells of the higher plants and like them they produce a definite number (four or eight) of daughter-cells, the spores, which contain haploid nuclei.
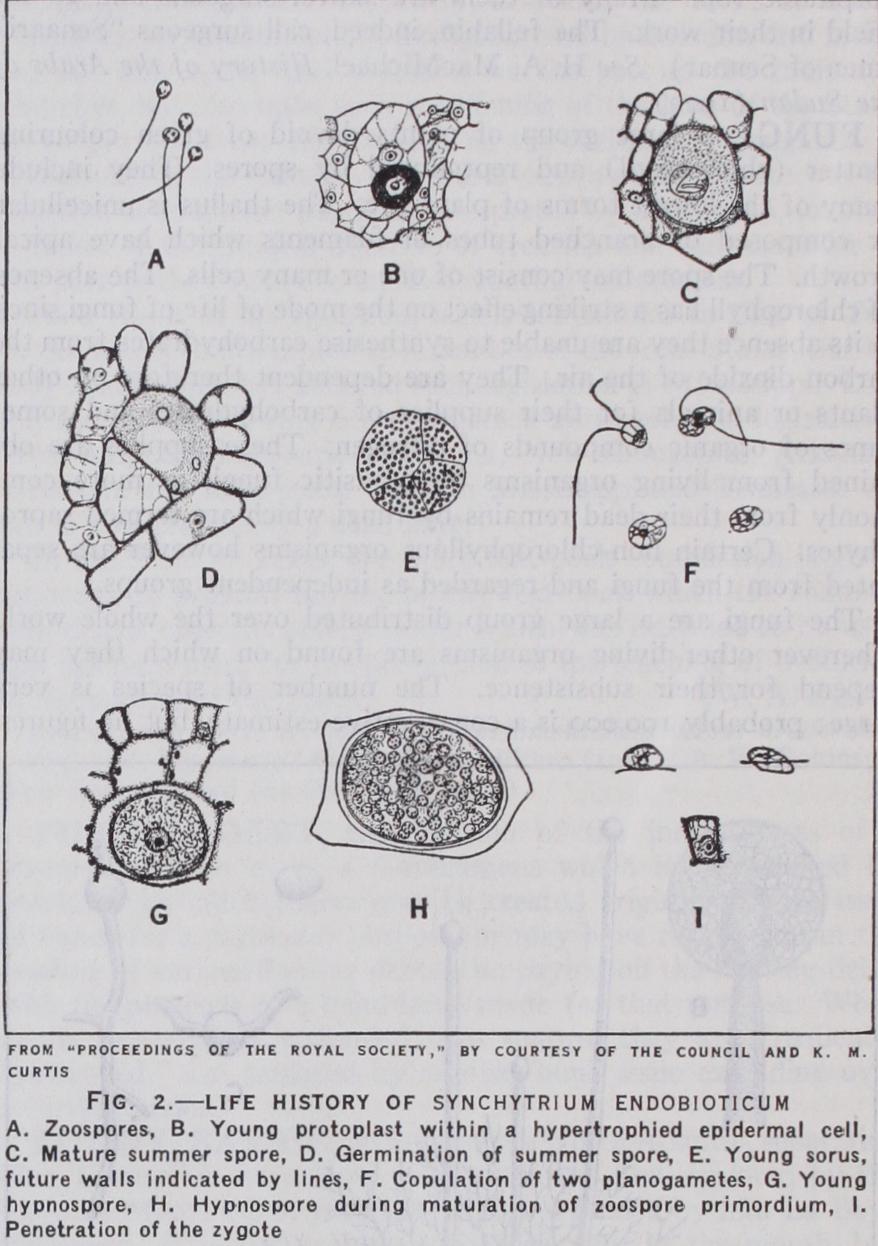
Life History of a Simple Fungus.—Some of the essential facts referred to above may be illustrated by reference to the life history of one of the simplest of the lower fungi, Synchytrium endobioticum, a parasite on potato tubers where it causes the well-known and destructive wart disease. The parasite enters an epidermal cell of the potato as a small naked protoplast con taining a single nucleus. It increases in size at the expense of the contents of the host cells and, when these are exhausted, its vegetative activity is brought to an end. It now surrounds itself with a wall of two layers, a thicker outer exospore and a thinner endospore and becomes a summer spore (fig. 2 c) . The exospore then ruptures and allows the endospore (prosorus), still enclosing the protoplast, to escape into the host cell. Here the nucleus begins to divide and by successive divisions, a number of nuclei, usually 32, are formed. The whole contents now become divided into several (five to seven) portions each of which becomes sur rounded by a separate wall. We have thus formed a group of cells in which as many as 30o nuclei may be formed by further division and the whole protoplasmic contents become cleft into as many small naked spores each containing a single nucleus. The cells containing these spores (the sporangia) burst and liberate their contents and each spore is seen to be provided with a motile organ in the form of a delicate protoplasmic thread or cilium. By the cilia the spores are able to propel themselves in the soil water and, from this animal-like movement, are known as zoospores. They settle down eventually on the surface of a young tuber, throw off their cilia and then penetrate the outer cell-wall of the host and so effect an entrance to the interior of the cell. Here they repeat the development described above. Thus the haploid generation of Synchytrium can reproduce itself and the cycle of zoospore-vegetative-cell-summer-spore-sporangia zoospore can be many times repeated. The release of the zoospores from the ripe sporangia is dependent on the presence of free water. If their escape is delayed for a time they show a signifi cant difference in behaviour. They swim towards other zoospores, they fuse together in pairs and the fusion cells, now provided with two cilia, after independent movement, settle down and infect the cells of the host (fig. 2 f, g). As many sporangia will liberate their contents simultaneously when wetted, it is probable that any two cells uniting together will be derived from different sporangia. The zoospores in this case behave as sexual cells or gametes; there is no morphological difference; a zoospore is a potential gamete and its behaviour is determined by physiological conditions. The young zygote contains two nuclei which fuse and form a diploid nucleus. In the host cell it enlarges and on the exhaustion of available food surrounds itself with a double wall. So far its development resembles that of a haploid plant, but it now shows a marked difference in behaviour, since it passes into a resting condition, which may persist until the next spring. It is therefore known as a resting spore (fig. 2 i). Eventually the nucleus divides and it is probable that there is a true meiotic or reduction division. After repeated divisions, zoospores are formed and set free by the splitting of the thick wall. Thus the haploid generation again begins its development. Here, then, we have the nuclear cycle completed, and the haploid and diploid phases are each associated with a distinct generation of the fungus which, though resembling one another in vegetative structure, differ in the final products of their development. The haploid generation is terminated by the production of gametes and their conjugation; the diploid generation is terminated by meiosis and the resulting production of haploid zoospores.
Fertilization.—The conjugation of two similar gametes (isogamy) is characteristic only of the simpler fungi. Through out the whole group it is possible to trace tendencies, which have not only profoundly modified the type of sexual reproduction, but in so doing have directly contributed to the success of the fungi in their transition from an aquatic to a terrestrial mode of life. These tendencies have led to the suppression of the individualiza tion of the gametes and the assumption of their functions by the gametangia—the structures in which gametes are formed.
In the simplest cases this has resulted in the sexual process consisting in the fusion of two entire individuals. These are usually of different size and we can distinguish a smaller male and a larger female (Polyphagus, fig. 3 d) . The mature individuals thus form a single gametangium and the distinction of sex is marked by calling the male gametangium an antheridium and the female an oogonium.
In most fungi the whole plant is not involved in reproduction as in Synchytrium and Polyphagus, but only certain hyphae which bear the gametangia. Thus many gametangia are borne on each individual and here we can distinguish two possibilities. The indi vidual may bear gametangia of both kinds (monoecious, herma phrodite, bisexual) or there may be two kinds of plants each bearing one kind of gametangium only (dioecious, unisexual). Since in the fungi we have only to consider sex in relation to the haploid generation, it is more convenient to distinguish these fungi by the terms first suggested by Blakeslee for the Mucor aceae—homothallic (monoecious) and heterothallic (dioecious) . Thus Polyphagus is heterothallic.
In the Mucoraceae in which both types of thallus occur, the gametangia, which contain a large number of nuclei representing the potential gametes, show no morphological distinctions into antheridium and oogonium. When they fuse their contents are mingled and a large number of nuclei pair together; we thus have multiple fertilization which, however, only results in the produc tion of one zygote, a coenozygote.
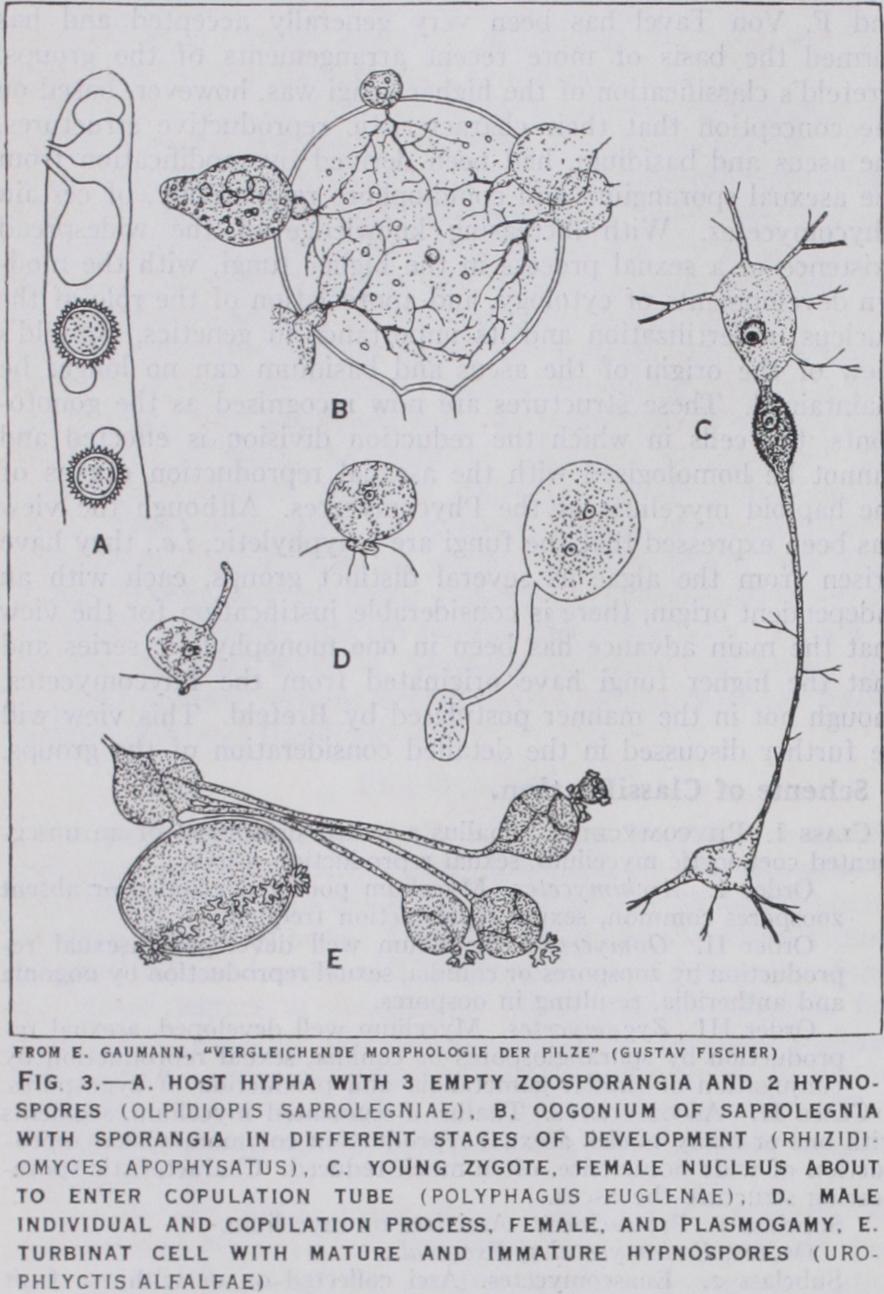
In the Oomycetes, homothallism is the rule (except Dictyuchus), and the gametangia are clearly differentiated into antheridia and oogonia. In Monoblepharis the antheridia give rise to a number of uniciliate male gametes which are set free in the water to find the oogonium. The latter's contents round off to form one large naked uninucleate mass termed the oosphere or egg (fig. 4 a ). This is not strictly a gamete, since it represents the whole con tents of the gametangium and at first contains many nuclei- potential gametes. In other members of this group (Saprolegnia ceae, Peronosporaceae) the element of chance in fertilization is still further reduced. The antheridia are brought into contact with the oogonia and the male gametes—now undifferentiated and represented only by their nuclei—are brought directly into contact with the eggs (fig. 4 d, e). As in the Mucoraceae, these fungi are independent of water for fertilization. It is this independence of water both for fertilization and, as will be shown later, for the dissemination of spores, that has rendered possible the migration of fungi from a life in water to an existence on land and on land plants.
In the higher fungi a further modification can be traced which leads to the suppression of the function of the male organ and finally to its disappearance. This does not involve the absence of fertilization, but the fusion of nuclei derived from nearly re lated cells of the female organ (Ascobolus) or of nuclei within the common mother-cell (Humaria, Lachnea). This leads finally to the disappearance of the female sexual organs and a condition of pseudogamy in which the cells concerned are two vegetative cells sexually different but not differentiated in structure. This is the case, e.g., in many yeasts and in some of the higher Basi diomycetes. These fusions of vegetative cells may appear to be on all fours with the anastomoses which occur not uncommonly between ordinary vegetative hyphae, but that they differ in kind is clear from the fact that they have the essential character of a sexual fusion inasmuch as they form the starting point of new development. As in all types of fertilization they mark the be ginning of the diploid generation. A reference to the conditions existing in Synchytrium, where every zoospore is a potential gamete, may help the realization, that in a haploid or gametophyte generation of one of the higher fungi, however numerous the cells or diverse in structure, every nucleus has the same organization, every cell has the potentialities of a sexual cell.
Asexual Reproduction.—The reproductive bodies of fungi are known as spores. The simplest type of reproduction is the separation of a cell or group of cells which is capable of inde pendent growth and which thus serves to propagate the fungus. Although physiologically these behave as spores they are usually distinguished as oidia (Mohilia) or, in sprouting fungi such as the yeasts, as buds or gemmae. From a morphological standpoint spores are marked off from the hyphae which produce them by their form, size, colour, mode and place of origin, etc. Spores formed within a special sac, the sporangium, are known as sporan giospores, or if motile as zoospores. If they are cut off terminally from a hypha they are conidia and the parent hypha is a conidiophore. Chlamydospores are thick-walled, usually resting spores formed like conidia or more usually as bodies on ordinary hyphae. The various kinds of spores of fungi also receive special names in the different groups, and these, although given without any reference to general principles, are useful and convenient owing to their descriptive character.
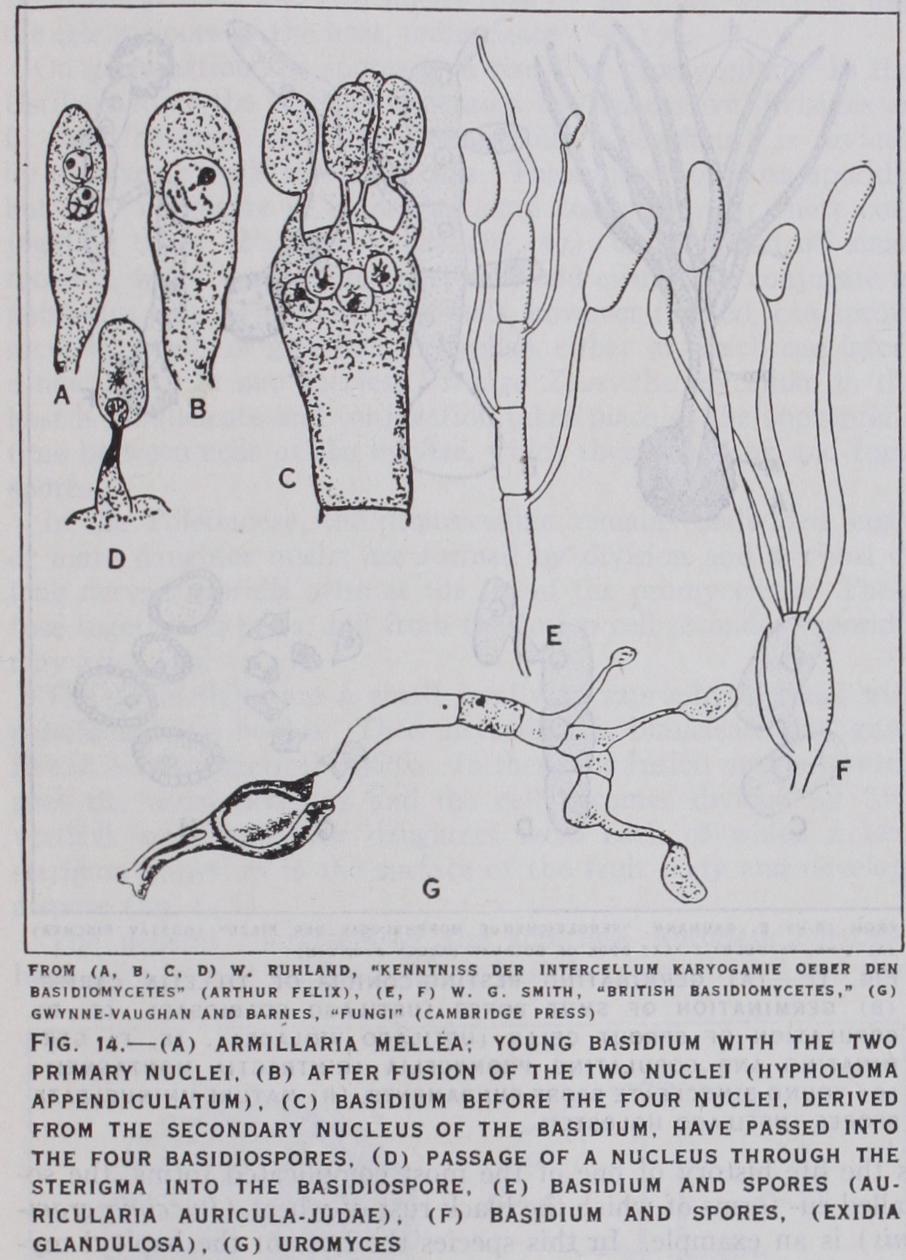
The various spore forms referred to above—sporangiospores, conidia, etc., are sometimes described as "summer" spores. It is a somewhat misleading term, but it does indicate that these bodies serve the purpose of rapid propagation of the fungus under favourable conditions. They are produced for the most part on haploid mycelia and appear as soon as the necessary food-materials have been accumulated by the vegetative hyphae. In the higher fungi special types of fruit bodies occur which are distinguished by the fact that they consist wholly or in part of diploid hyphae and that they are concerned with the production of gonotoconts, those cells in which the reduction divisions are effected. The gonotoconts produce daughter cells—the spores—with haploid nuclei. Where the spores are produced within the mother-cell endogenously—it is called an ascus and the daughter cells asco spores. Where they are borne externally—exogenously—they are termed basidiospores and the mother cell a basidium. Details of the structure and mode of formation of these fruit bodies will be given in later sections dealing with classification (see below, Ascomycetes and Basidiomycetes).
The general physiology of the fungi does not differ essentially from that of other plants. Their requirements are the same as those of the living cells devoid of chlorophyll of the higher plants, wherever these may occur. Like them they require water, various inorganic salts and supplies of carbonaceous and nitrogenous foods in varying degrees of complexity. They respire, taking in free oxygen and liberating free carbon dioxide or excreting carbonic acid. Aerial hyphae transpire and often exude drops of water; waste products (e.g., calcium oxalate) are also excreted. They respond to external stimuli such as light, gravity, certain chemical substances, etc.
The one fact which dominates and restricts the mode of life and nutrition of the fungi is the absence of chlorophyll and their consequent dependence on outside supplies of organic carbon and in many cases of organic nitrogen. Fungi are therefore com pelled to live on materials derived from other plants or from animals, and are either parasites on living organisms or sapro phytes living on their dead remains. No hard and fast line can be drawn between these habits. There are many fungi normally living as parasites which can exist as saprophytes and vice versa. The large majority of fungi are saprophytes and they, with the bacteria, are the great agents of decay in nature. The vast amount of vegetable material which annually falls to the soil, the remains of herbs, the fallen leaves of deciduous trees, the fallen branches and trunks of forest giants, the excrement and the dead bodies of animals are all material for the sustenance of fungi. They are abundant in the upper layers of the soil in which these organic remains become incorporated by the action of many animals, e.g., worms, beetles, etc. In the household they are found on stale foods of every kind, on preserves, on damp clothing and leather, wall paper and plaster. Special means are therefore taken to protect foods from destruction by the common moulds.
The whole question of the significance of these structures and of the relations existing between the fungus and its host, has provided a wide field for speculation and in recent years has led to experimental investigations which have yielded important and suggestive results. On the whole, these investigations may be said to support the view that these structures are of benefit to the host plant. It does not follow that any obligate relation exists between the fungus and the host. In the case of forest trees seedlings can be grown successfully in the absence of the fungus, but there is strong evidence that the presence of the fungus may promote the absorption of nitrogenous substances from the humus of acid soils and this may well be a decisive factor in the success of trees established in such conditions. In the case of many orchids whose seeds are notoriously difficult to germinate, suc cessful growth can be induced by sowing the seeds on a suitable medium containing the appropriate endophytic fungus. Even here, however, the same results can be obtained in the absence of the fungus by a careful adjustment of the composition of the medium. Interesting as these latter results may be, as indicating the possible role of the fungus in promoting germination, it can hardly be doubted that in nature the latter plays a decisive part. It is probable that in these plants, and in forest trees, the fungus in the root cortex may serve to assist in the absorption of organic compounds. The strong development of similar fungi in the tissues of those orchids, which are devoid of chlorophyll and must therefore depend for their whole supply of organic food on the humus in which they live, gives strong support to this conclusion. There is no evidence that the plant in these cases is able to undertake this absorption in the absence of the fungus. The presence of fungi, which probably have a similar significance to those of mycorrhizas, has been noted in other groups of plants. They are known to be of regular occurrence in many liverworts, the prothallia of Lycopodium, the rhizome of Psilotum and in a number of ferns (Ophioglossaceae, Marattiaceae) .
In addition to these forms, many other fungi contain poisonous substances such as alkaloids which render them extremely danger ous when used as foods (species of Amanita, Stropharia, Russula, Boletus, etc. See Plate). However, there are a number of fungi which are perfectly wholesome when properly prepared, and although in England the common mushroom (Psalliota cam pestris) alone is generally eaten, on the continent of Europe many other species are eagerly sought, e.g., Lepiota procera, Morchella esculenta, Boletus edulis, Cantharellus cibarius. In addition to these species the truffle (Tuber) is universally esteemed a great delicacy.
The fungi which are the most important from an economic standpoint are the yeasts. They owe their importance to their power of fermenting various sugars with the production of alcohol (see BREWING). Brewers' yeast is also used in bread-making, the "rising" of the bread being caused by the escape of the bubbles of carbon dioxide evolved by the fermenting yeast. Another fungus, Mycoderma aceti, can, in the presence of oxygen, change the alcohol of fermented liquors to acetic acid and so produce vinegars. Of minor importance may be mentioned the species of Penicillium with which various cheeses (Stilton, Gorgonzola) are inoculated in order to produce the characteristic green-veining and improvement in flavour.
It has for long been recognised that the fungi proper fall into three main groups, the Phycomycetes, Ascomycetes and Basidio mycetes. The scheme of classification put forward by O. Brefeld and F. Von Tavel has been very generally accepted and has formed the basis of more recent arrangements of the groups. Brefeld's classification of the higher fungi was, however, based on the conception that their characteristic reproductive structures, the ascus and basidium, had been derived by modification from the asexual sporangium and conidiophore respectively, of certain Phycomycetes. With increasing knowledge of the widespread existence of a sexual process in the higher fungi, with the mod ern developments of cytology, and appreciation of the role of the nucleus in fertilization and its importance in genetics, Brefeld's view of the origin of the ascus and basidium can no longer be maintained. These structures are now recognised as the gonoto conts, the cells in which the reduction division is effected and cannot be homologised with the asexual reproduction organs of the haploid mycelium of the Phycomycetes. Although the view has been expressed that the fungi are polyphyletic, i.e., they have arisen from the algae as several distinct groups, each with an independent origin, there is considerable justification for the view that the main advance has been in one monophyletic series and that the higher fungi have originated from the Phycomycetes, though not in the manner postulated by Brefeld. This view will be further discussed in the detailed consideration of the groups.
Scheme of Classification.
Phycomycetes.—I. The Archimycetes. These parasitic and mainly aquatic forms have been looked upon as degenerate Oomy cetes and were formerly included in that group. Simplicity of structure, however, though it is often the result of degradation, is equally a mark of primitive organisation. The fact that in this group is found the simplest type of sexual reproduction known among the fungi is justification for regarding them as an assem blage of primitive organisms, whose relation to the other groups of Phycomycetes is somewhat obscure. The simplest representatives such as 0lpidium and Synchytrium live in the cells of their hosts as naked protoplasts. They reproduce by uniciliate zoospores, which under certain conditions behave as gametes (see above Synchytrium) . The zygote resulting from the conjugation of the gametes has a period of vegetative growth and encysts to form a resting spore. A similar mode of life and vegetative structure is exhibited by Olpidiopsis in which the zoospores are biciliate; here, however, there are no motile gametes, a larger female and a smaller male individual fusing together. These are multinucleate and may be regarded as gametangia which have assumed the func tions of gametes (fig. 3 a).
In Rhizopliidium the parasite is enclosed in a membrane and lives outside its host, into which it sends a short peg-like hausto rium, which in some related forms (Rliizidiomyces), develops delicate absorbing threads (fig. 3 b). At maturity the whole body becomes a sporangium or acquires a thick wall and forms a resting spore. Sexual reproduction is unknown, but in the re lated Zygorhizidium conjugation between two individuals creates a resting zygote (fig. 3 d). This mode of sexual reproduction in which the conjugation of two individual plants is concerned is found in other forms, e.g., Sporophlyctis, Polyphagus. In the latter, which is parasitic on Euglena, the fungus possesses a highly developed system of hyphal-like haustorial threads, enabling it to attack a number of alga cells simultaneously (fig. 3 c). In other members of the group these threads, though extremely deli cate, may be regarded as fine hyphae. In Physodema and Uro phlyctis they develop at intervals large swollen vesicles, the col lecting cells, which not only serve as centres for fresh hyphal development, but give rise to large, thick-walled resting spores (fig. 3 e).
Olpidium Brassicae causes a disease of cabbage seedlings, the species of Synchytrium cause warts or tumour-like swellings on their hosts (e.g., potato, dog's mercury, dandelion) and similar lesions are caused by species of Urophlyctis on alfalfa, beet and other plants.
II. The Oomycetes. These fungi have a well developed my celium of large coenocytic hyphae. The lower families (Mono blepharidaceae, Saprolegniaceae) are saprophytes, living in water on vegetable or animal remains, the Ancylistineae being reduced forms, parasitic on algae, eelworms. etc., the Peronosporaceae are saprophytes or highly specialised parasites on land plants.
The group exhibits in the structure both of its asexual and sexual organs a series of modifications, which can be directly re lated to the gradual change of habit from an aquatic to a terres trial existence.
The lower families are propagated by zoospores, uniciliate in Monoblepharis, biciliate in the Saprolegniaceae and Ancylistineae (fig 4 b) borne in sporangia which, e.g., in Saprolegnia, are but slightly enlarged terminal portions of the hyphae which bear them. In Pythium and Uiytophthora the sporangia are spherical or oval in form and in certain conditions (absence of free water) they can put out a germ tube and so give rise directly to a hypha. In other genera (Peronospora) this direct mode of germination is alone possible, zoospores are no longer produced, the zoosporan gium has become a conidium—capable of germinating in moist air and so adapted for a parasitic life on land plants (fig. 4 b) .
In their sexual reproduction the group exhibits very strikingly the tendency already mentioned to replace the union of gametes by the fusion of gametangia. In Monoblepharis the female organ, the oogonium, contains a single, non-motile, uninucleate egg. It is fertilized by a motile spermatozoid, which closely resembles in structure the zoospores. The zygote resulting surrounds itself with a thick membrane and forms a resting oospore (fig. 4 a). In Saprolegnia the young oogonium contains many nuclei, potential gametes, but of these only a few become each the nucleus of an egg, of which there are several in the mature oogonium. There are no spermatozoids, and male gametes are not differentiated, the antheridium puts forth a fertilization tube which conveys the male nuclei to the eggs. Thus, even in members of a family exclusively aquatic, water is no longer the means by which the male gametes reach the eggs. Some members of this family have but one egg in the oogonium and this is the rule in the other families. In Albugo Bliti the egg has many nuclei and numerous male nuclei pass from the antheridium and fuse with them. We have thus multiple fertilizations and the oospore which results may be re garded as a coenozygote since it contains many zygote nuclei, each the result of a separate fertilization. In most of the higher Oomy cetes however the mature egg is uninucleate and only one male nucleus passes into it from the fertilization tube. Thus the final result of the assumption of the function of the gametes by the gametangia is a single fertilization, made, however, under such conditions as to eliminate accidents and ensure the production of the zygote.
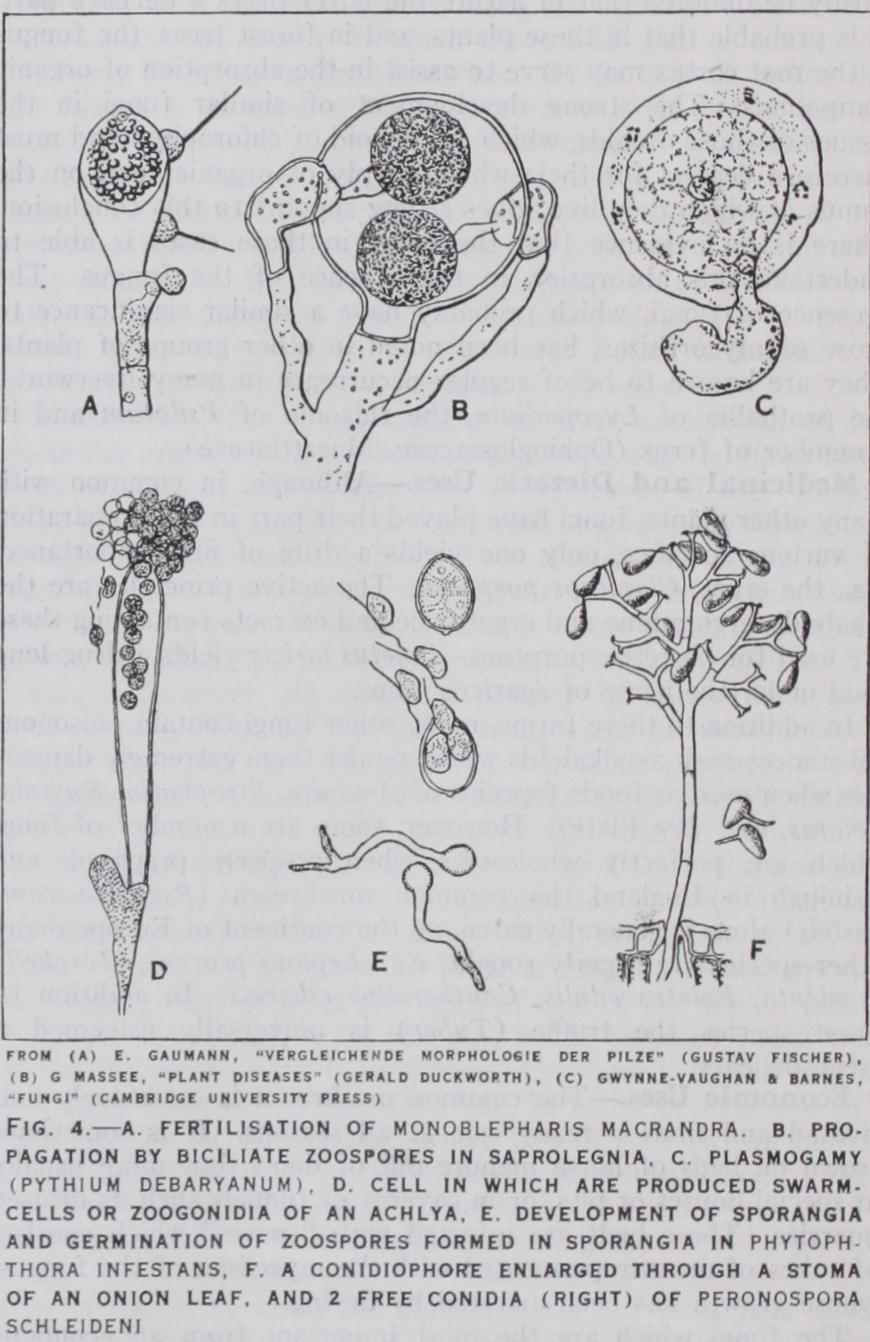
Many of this family are dangerous parasites of cultivated plants and the cause of destructive diseases, e.g. "damping off" disease of seedlings (Pythium), potato disease (Phytophthora), vine mildew (Plasmopara).
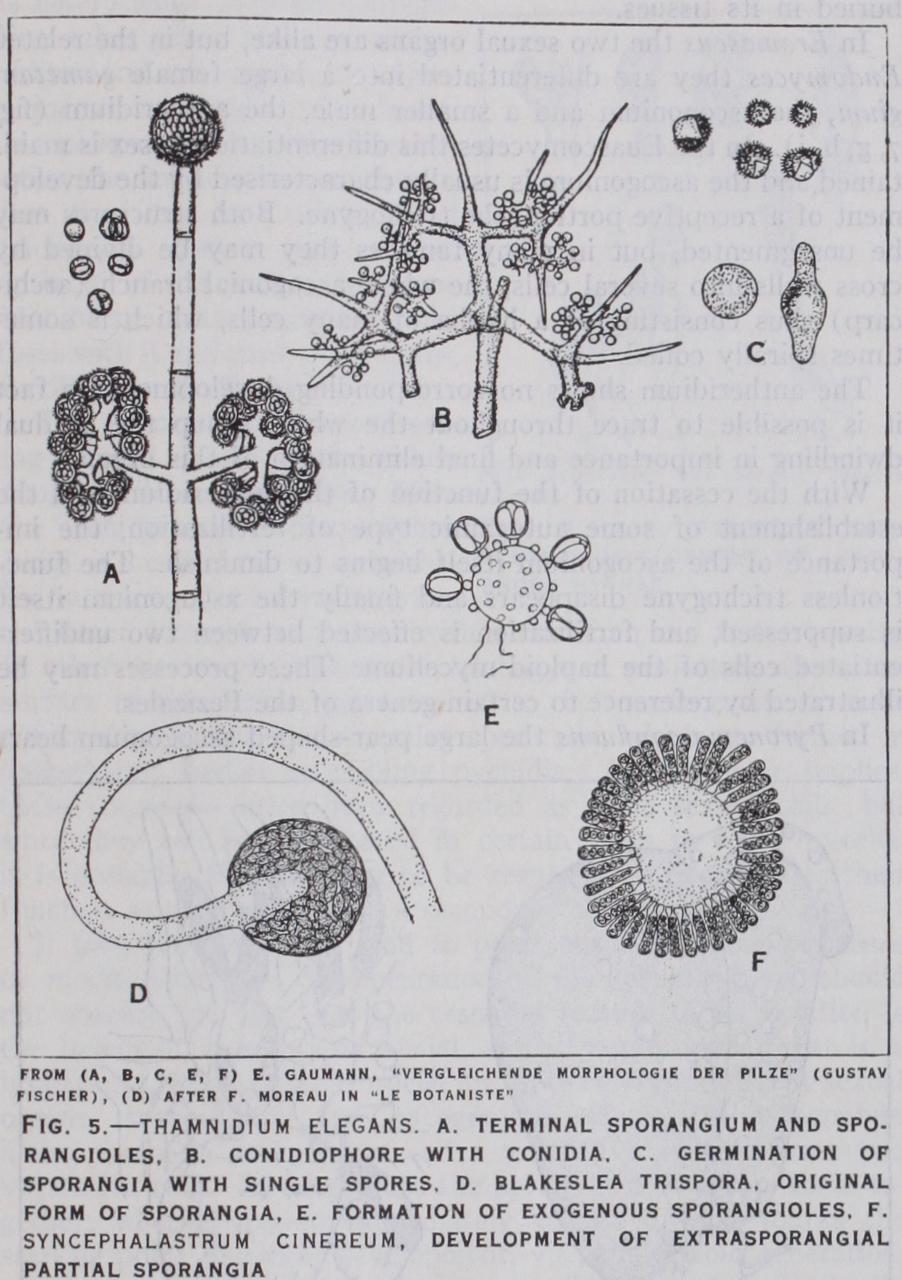
III. The Zygomycetes. These fungi are largely saprophytic in Iii. The Zygomycetes. These fungi are largely saprophytic in habit and terrestrial. Their mycelium resembles in structure that of the Oomycetes. Zoospores are unknown, the characteristic reproductive organs of the leading family, the Mucoraceae, being sporangia which may contain (Mucor) thousands of spores (fig. I b). Certain genera, however, are reproduced by conidia, e.g., Chaetocladium, Cunninghamella, and there are a series of forms which show that in this family, as in the Oomycetes, a transforma tion of sporangia to conidia has taken place. In one series this has been brought about by the gradual reduction in the number of spores in the sporangium; so, e.g., Thamnidium has small sporangia with two to four or sometimes only one spore and Chaetocladium may be regarded as a genus in which the small sporangia each contain a single spore and hence behave as conidia (fig. 5 a, b, c).
In another series, Blakeslea produces sporangia of normal type and also spherical heads bearing a number of small sporangia with only 3 spores. These can be related to the spherical conidiophores of Cunninghamella and Syncephalis.
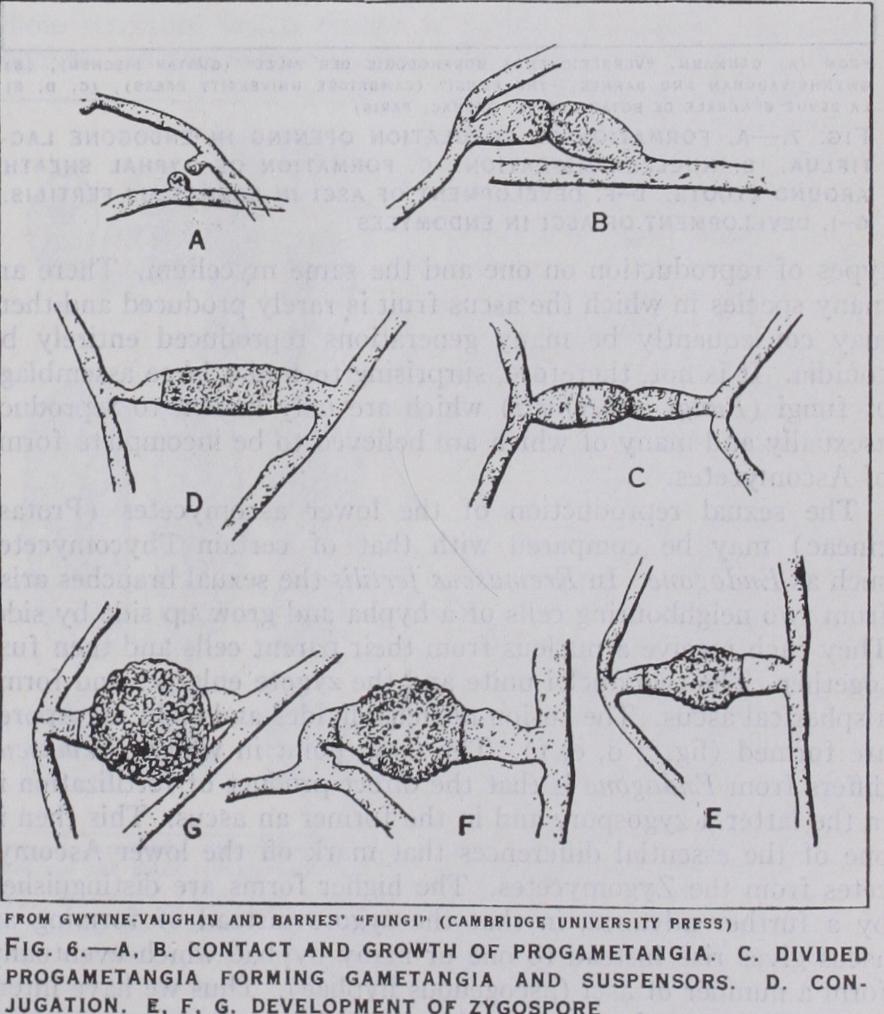
In their sexual reproduction the Mucoraceae exhibit the con jugation of coenocytic gametangia, the resulting structure, the zygospore, containing many zygote nuclei. The gametangia con cerned may arise on the same mycelium, on a common supporting hypha or zygophore as in Sporodinia, Zygorhynchus, and such forms are described as homothallic. In the majority of the species so far investigated the gametangia arise on two separate mycelia, which are different in kind. There is little doubt that these two kinds of mycelium represent distinct sexes. A. F. Blakeslee, to whom the discovery of this condition—known as heterothallism is due, pointed out that a difference of vigour, indicated, e.g., by a greater luxuriance of growth, was often to be observed between the two strains, which for this reason he distinguished as (+) and (—) . He was able to show, moreover, by the production of so-called hybrid reactions that there is an essential sameness in all the (+) and in all the (—) strains, respectively, through out the family, and that the (+) strains are probably to be regarded as female and the (—) as male.
The germination of the zygospore usually results in the produc tion of a "germ" sporangium and in Phycomyces nitens it is in this structure that the zygote nuclei undergo the reduction divi sion. The resulting spores are in the main of two kinds, producing (+) and (—) mycelia respectively; but there are also a few spores which give rise to feeble thalli—neutral mycelia—which are homothallic. It is of interest to note that H. Burgeff experi mentally produced similar mycelia by the admixture of the con tents of hyphae from (±) and (—) mycelia, and thus reached the conclusion that homothallic mycelia contain both (+) and (—) nuclei.
The Entomophthoraceae is a family of insect parasites of which one, Empusa ?miscue, attacks the common house-fly. They are reproduced asexually by conidia which are shot off from the conidiophores and form a halo around the dead flies.
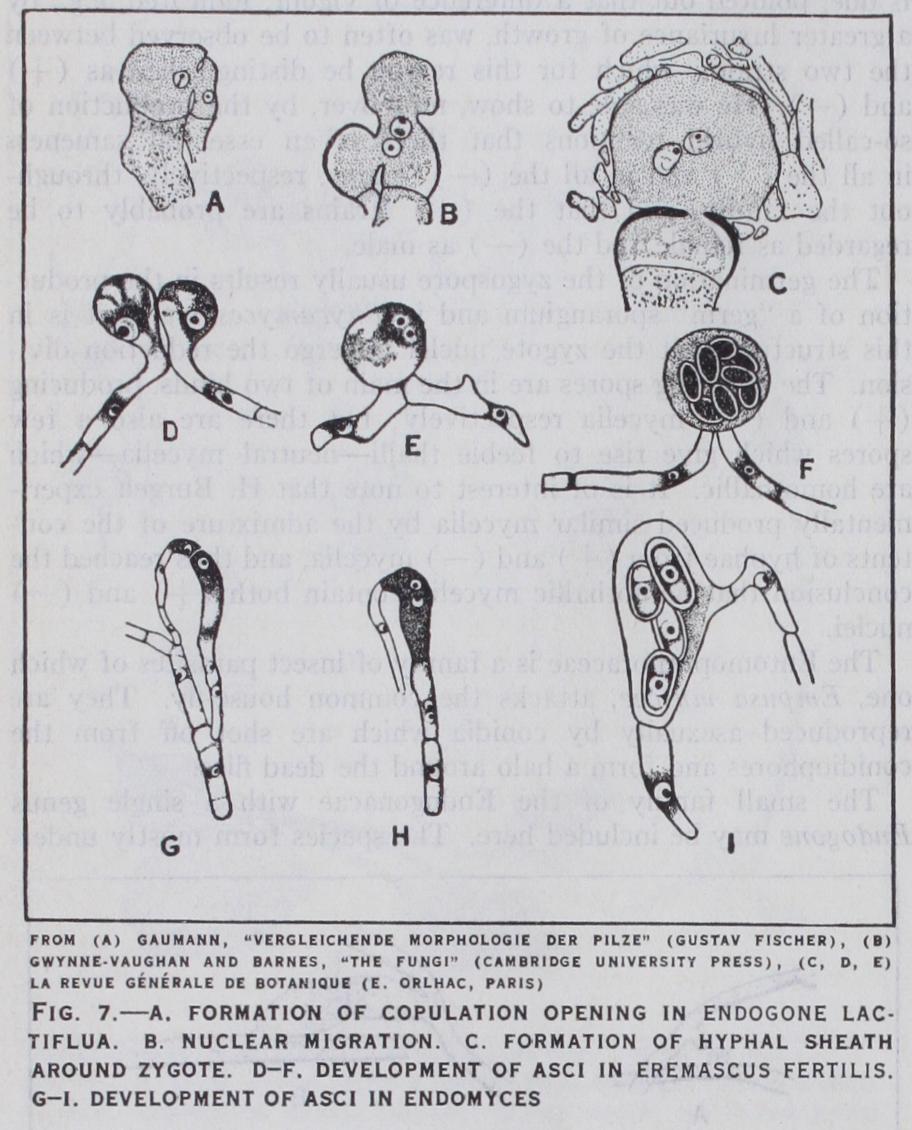
The small family of the Endogonacae with a single genus Endogone may be included here. The species form mostly under ground stromata in which the reproductive organs are embedded. In Endogone lactiflua two sexual branches of unequal size—male and female—grow up side by side. They are multinucleate at first and from each a terminal cell containing a single nucleus is cut off. These cells unite together and from the larger, the female, there now arises an outgrowth which rapidly enlarges, and into this pass the two sexual nuclei. This outgrowth is the zygote, which not only develops a thick wall but becomes surrounded and embedded by a number of vegetative hyphae (fig. 7 a, b, c).
Its germination is unknown. Endogone has been included in the Ascomycetes and is of considerable interest as indicating a possible relation between that class and the Zygomycetes.
The sexual reproduction of the lower ascomycetes (Protas cineae) may be compared with that of certain Phycomycetes such as Endogone. In Eremascus f ertilis the sexual branches arise from two neighbouring cells of a hypha and grow up side by side. They each receive a nucleus from their parent cells and then fuse together. The two nuclei unite and the zygote enlarges and forms a spherical ascus. The fusion nucleus divides and eight ascospores are formed (fig. 7 d, e, f). The main point in which Eremascus differs from Endogone is that the direct product of fertilization is in the latter a zygospore and in the former an ascus. This then is one of the essential differences that mark off the lower Ascomy cetes from the Zygomycetes. The higher forms are distinguished by a further advance, in that the zygote instead of forming an ascus gives rise instead to one or many hyphae which eventually form a number of asci (ascogenous hyphae). Thus we have inter calated between the gametangium and the ascus a new mycelial generation—the diploid generation.
The development of this diploid generation is accompanied in many forms with a delay in the fusion of the sexual nuclei, so that the ascogenous hyphae contain paired nuclei (dikaryons). It is in the ascus that nuclear fusion is finally accomplished, to be succeeded immediately by the meiotic phase and the formation of spores.
The ascogenous hyphae, however, do not form an independent self-supporting mycelium. They are dependent on the haploid mycelium for their nutrition, and this is accomplished by the enclosure of the ascogenous hyphae in a simultaneous growth of vegetative hyphae from the parent mycelium. Thus we have formed the fruit body or ascophore, one of the characteristic structures of the higher ascomycetes, showing in the different families great variety in structure and organisation. Whilst in many families its development awaits the stimulus which origi nates in fertilization, in the higher forms it may reach an ad vanced stage of development before the sexual organs arise, buried in its tissues.
In Eremascus the two sexual organs are alike, but in the related Endomyces they are differentiated into a large female gametan gium, the ascogonium and a smaller male, the antheridium (fig. 7, g, h, i). In the Euascomycetes this differentiation in sex is main tained and the ascogonium is usually characterised by the develop ment of a receptive portion, the trichogyne. Both structures may be unsegmented, but in many families they may be divided by cross walls into several cells, the whole ascogonial branch (archi carp) thus consisting of a hypha of many cells, which is some times spirally coiled.
The antheridium shows no corresponding development, in fact it is possible to trace throughout the whole group the gradual dwindling in importance and final elimination of this organ.
With the cessation of the function of the antheridium and the establishment of some autogamic type of fertilization, the im portance of the ascogonium itself begins to diminish. The func tionless trichogyne disappears and finally the ascogonium itself is suppressed, and fertilization is effected between two undiffer entiated cells of the haploid mycelium. These processes may be illustrated by reference to certain genera of the Pezizales.
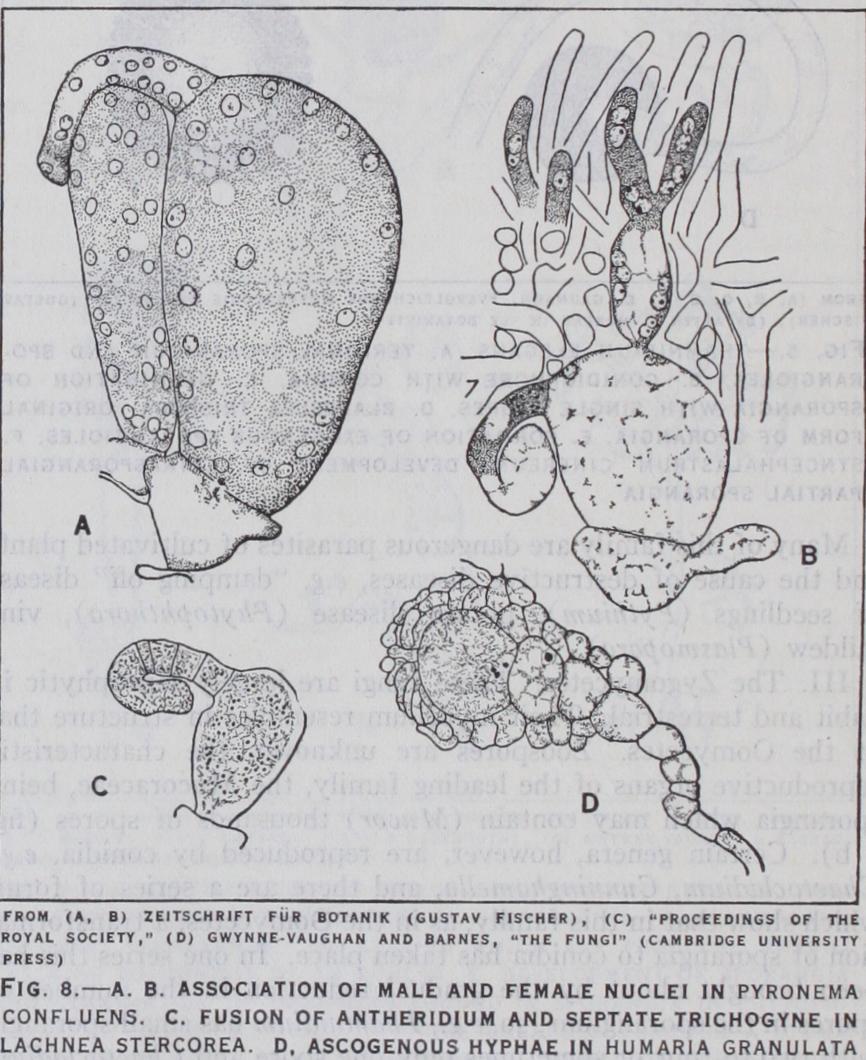
In Pyronema confluens the large pear-shaped ascogonium bears at its upper end a tubular trichogyne, the tip of which fuses with the club-shaped antheridium. All these structures are multinucle ate. The contents of the antheridium pass first into the trichogyne and then, by absorption of the intervening wall, into the as cogonium, where the male and female nuclei become associated (fig. 8 a, b). In Lachnea stercorea, though the antheridium is fused with the trichogyne, the latter is septate and no male nuclei pass from the antheridium into the trichogyne (fig. 8 c). In Lachnea cretea no antheridium is formed though the septate tri chogyne is still developed. In Humaria granulata the trichogyne has disappeared though the ascogonium has still the form of those described above and like them gives origin to the ascogenous hyphae (fig. 8 d). Finally in Humaria rutilans the ascogonium itself has disappeared and, with it, all trace of autogamous fertili zation, which is replaced by the pseudogamous fusion of vegetative cells.
In one species Ascobolus carbon, arias the antheridia are absent, but the long and often coiled trichogyne comes into contact with one of the oval, stalked con idia, borne on the mycelium, and fuses with it and thus, apparently, fertilization is effected.
This process finds an interest ing parallel in one of the lichen fungi Collema pulposum. Here the archicarp, as in many lichen fungi, is a coiled thread terminat ing in a long slender trichogyne; the latter comes into contact with a conidium and fusion results. In other species of Collema the trichogynes, projecting above the surface of the thallus, get attached to their surface one or more uninucleate bodies, the so called spermatia, which are formed in flask-shaped bodies resembling pycnidia. As the name implies, these spermatia have been regarded as male sexual cells, but since they can be germinated in certain cases to form mycelia, it is possible that they are to be regarded as pycnospores their function as male cells being a secondary one.
It may perhaps be as well to point out, that these processes of modification and disappearance of the sexual organs, should not obscure the fact that the essential feature of fertilization is the fusion of two haploid nuclei, and it would appear that it is immaterial, whether these nuclei are derived from different sexual organs, antheridium and ascogonium, from the ascogonium alone, from an ascogonium and a vegetative cell, or from two vegetative cells. In every case the process which brings them to gether, whether fusion immediately results or not, marks the starting point of a new development, viz., the diploid generation, represented by the ascogenous hyphae. These hyphae eventually give rise to the asci and in the majority of the ascomycetes they are formed in a very characteristic way. As already stated the ultimate segments of these hyphae are binucleate and any one of them may grow out to a short branch, into which the two nuclei pass. This cell now curves over at the tip to form a hook, and the nuclei moving up to the summit divide there simultaneously. The spindles are so arranged that when the two cross walls are formed the summit of the crook contains two nuclei, whilst one is in the tip and another in the stalk (fig. 9).
In most cases the two nuclei at the summit fuse together and form the fusion nucleus of an ascus, which is formed by the further development of the parent cell. The point of the crook and the stalk lie close together and either of them may now become binucleate by the migration into it of the nucleus from the other. In this process the walls are perforated by the travel ling nucleus. The binucleate cell so formed may now originate a new crook and so the process may be repeated several times.
The young ascus now enlarges, the nucleus also increasing in size, and finally the latter divides; this is followed by two further divisions and thus eight nuclei are formed, which become the centres in forming eight ascospores (fig. io). The details of nuclear division in the ascus have been carefully studied in a number of forms, and there is no doubt that the first division in the ascus is the heterotype or reducing division (see CYTOLOGY). The two following divisions have, for the most part, the characters of post meiotic divisions, but there is no general agreement as to their interpretation. In the account of the sexual process and development of the ascogenous hyphae already given, it has been assumed that the sexual nuclei are associated in pairs and pass as such (dikaryons) into the ascogenous hyphae; here the pairs may divide but later a pair of nuclei, by derivation male and female, fuse together in the ascus and so complete fertilization. In such a case the first division in the ascus is a reducing division and in the two succeeding divisions the haploid numbers of chromosomes will appear.
There are a number of forms however (Sphaerotheca, Humaria, Lachnea, Ascobolus) in which the nuclei in the ascogonium, how ever derived, have been described as fusing together. In this case, the nuclei in the ascogenous hyphae are diploid nuclei and the fusion in the ascus is a second fusion, and results in a nucleus with a fourfold number of chromosomes (tetraploid nucleus) . Obviously a double process of reduction is necessary to restore the haploid nuclei of the sexual generation, and the second reduc tion is stated to take place during the second or third division. Further investigation is needed either to confirm or disprove the existence of double fusion and double reduction.
The weight of evidence on the whole favours the conclusion, that single fusion is the normal process in the Ascomycetes. In this respect they conform with the other large group of the higher fungi, the Basidiomycetes.
The mode of spore-formation in the ascus is peculiar. The eight nuclei formed in the third division show delicate radiations, extending from a centrosome, which meet around the nucleus and thus delimit a portion of the protoplasm on which a wall is secreted to form a young spore (fig. 1o, h, j) .
It has already been explained that, in the higher Ascomycetes, the developing ascogenous hyphae are enclosed by a development of the haploid mycelium, on which, in fact, they are parasitic. The whole structure which results is known as a fruit body or as cophore. In the simpler orders (Plectascales and Perisporiales) the enclosure is complete, the sterile hyphae compacting to gether on the outside to form a completely closed investment. Such a fruit body is called a pera thecium. In other orders (Hy pocreales, Sphaeriales, Dothid eales) a perithecium is formed, which is often flask-shaped, and provided with a neck with a definite opening for the discharge of the spores (fig. 12).
The remaining orders have a fruit-body known as an apothe cium. Here the numerous asci are assembled, together with sterile hyphae or paraphyses, in a continuous layer known as the hymenium (fig. 13) . This is freely exposed on the surface of the ripe fruit body which may be a flat disc or a (Pezizaceae) or a convex, often stalked, struc ture (Helvellaceae).
The spores of many ascomy cetes are explosively discharged from the asci ; a process which is facilitated by their mucilaginous contents and the pressure of the surrounding asci and paraphyses.
When spores are sown in suitable culture fluids they bud after the manner of yeast cells and produce conidia. In two species of Taptirena it has been shown that these secondary conidia can con jugate in pairs by means of a short tubular process. The nucleus from one cell passes into the other and the binucleate cell so formed now puts out a germ-tube. Moreover, when the eight spores from one ascus are carefully isolated and germinated apart, it is found that they fall into two groups, the conidia of one group will not conjugate together but do so readily with any member of the other group. They are thus, like many of the Muco rini, heterothallic, and the separation into the two sexes must take place during the divisions in the ascus.
The Plectascales include the common saprophytic moulds Aspergillus and Penicillium of which some species—Aspergillus herbariorum, Penicillium crustaceum are the commonest and most ubiquitous of fungi. They reproduce themselves by a profuse pro duction of conidia and also by closed (cleistocarpous) perithecia (fig. II).
The Perisporiales are parasitic fungi living epiphytically, i.e., on the surface of their hosts, absorbing food materials by haus toria from the epidermal cells—or sending hyphae through the pores of the stomata into the interior of the host. The Erysipha ceae or mildews are the cause of disease in many cultivated plants, e.g., hop mildew, American gooseberry mildew. They produce conidia in chains and numerous perithecia which bear character istic appendages. The Perisporiaceae with a similar habit have dark coloured mycelium and are known as sooty moulds.
The three orders Hypocreales, Sphaeriales, Dothideales include families which have frequently been grouped together under the names of Pyrenomycetes. They have perithecia which, when ma ture, have a well-defined mouth (fig. 12). In some genera the fruit bodies are borne directly on the mycelium (Sordaria, tomium) or isolated in the host tissues (Venturia) but more often they are borne on a stroma tria, Cucurbitaria) or embedded in it (Epichloe,Claviceps,Valsa). The stroma may be flat or ion-shaped, or immersed in the tissues of the host. In the more specialized forms the stroma may show a sterile basal stalk, the perithecia being limited to the upper part, where they may more efficiently distribute their spores (Xylaria). The mycetes is by far the largest group of the Ascomycetes, a servative estimate of the ber of species being I o,000. Though the majority are phytes some well-known diseases of plants are caused by members of this group, e.g., canker in fruit trees (Nectria), ergot of cereals and other grasses (Claviceps), apple and pear scab (Venturi˘), most species of Cordiceps are parasites on insects.
The Pezizales include the forms with apothecia and they pro vide the largest and most elaborate fruit-bodies among the as comycetes, e.g., Morchella which may be io–i 2 inches in height.
In the Pezizaceae—the cup fungi—the apothecium is con cave, saucer or cup-shaped, sometimes stalked. In the Geoglossa ceae and Helvellaceae the ascophores are club-shaped or differ entiated into a sterile stalk and fertile head, which may be folded or ridged in various ways to increase the area of the spore-bearing surface (fig. 13).
The Tuberales is a small group of fungi which include the well known truffles (Tuber sp.). They live underground in the neigh bourhood of trees and have been thought to be concerned in the formation of mycorrhizas. The hymenium lines internal chambers in the fruit-bodies.
The Laboulbeniales is an order of peculiar fungi, whose mani fold structure has been made known by the investigations of Thaxter in America. They are minute plants from I-10 m.m. in height which grow attached to the bodies of insects, especially beetles. They are dark brown in colour and consist of a basal receptacle which bears appendages, on which are the sexual organs. The female organ is essentially an ascogonium with a trichogyne and is fertilized by male cells or spermatia. The fertilized ascogenic cell (ascogonium) gives rise to a number of asci which are enclosed in a perithecial wall that develops around the female organ. Some species are probably heterothallic (Amorphomyces) but the male and female spores are associated together in the ascus, are shed and germinate together.
Though sexual organs are absent the haploid mycelium is often sexually differentiated. Heterothallism occurs both in the Ure dinales and Ustilaginales, and is widespread in the higher forms. In the latter, pseudogamous fusions presumably occur between hyphae of the mycelia of different kinds, though in only a few cases have these actually been observed. Asexual reproduction by conidia and oidia occurs in both types of mycelia.
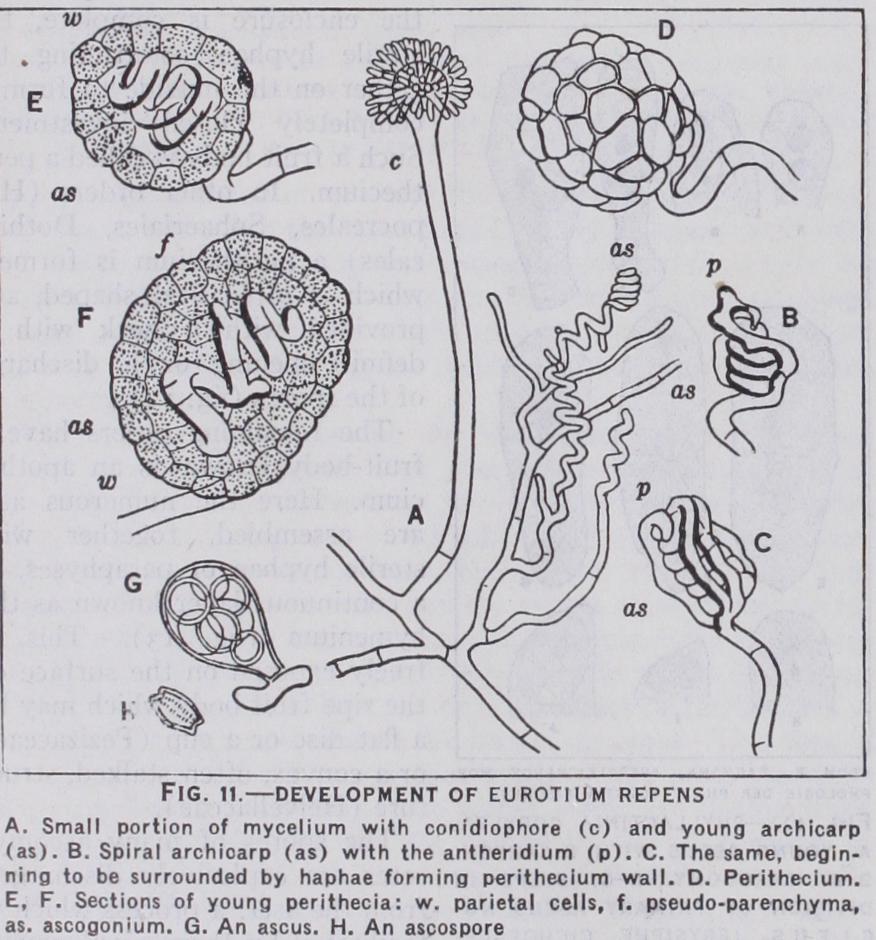
In the lower families the basidia are borne directly on the diploid mycelium, but in the vast majority of the forms they are borne on fruit bodies or sporophores. These are not formed, as in most Ascomycetes, as the immediate result of a sexual process (see above) but arise from the diploid mycelium at a time and stage in its development which are determined by physiological factors, such as nutritive conditions, humidity, light, etc. The basidium, like the ascus, is the gonotocont. In it the reduction of the chromosomes is effected but the daughter cells (usually four) resulting are exogenous, i.e., borne on short stalks (sterig mata) outside the mother cell (fig. 14 d). Two kinds of basidium occur, those characteristic of the Autobasidiomycetes are, like the ascus, undivided (holobasidia). The Protobasidiomycetes have the basidia divided by septa into four cells (phragmobasidia) (fig. 14 e). The dividing walls may run transversely, e.g., Auricularia, or longitudinally as in Trernella (fig. 14 f). The fertilization which takes place in the young basidium is followed immediately by the reducing divisions and the development of spores, but in the Uredinales and Ustilaginales, after the nuclear fusion, a resting stage, often very prolonged, may intervene. This may be accom panied by a thickening of the wall of the mother-cell, which may even become set free from the mycelium as a spore (teleutospore, chlamydospore). These "probasidia" as they may be called, can no longer behave as basidia and produce the spores; they conse quently "germinate," that is they produce a short vegetative out growth resembling a germ tube (promycelium), which functions as the basidium and bears the basidiospores (sporidia) Thus in these forms the probasidium is concerned with the distribution and perennation of the fungus as well as with the production of the basidiospores (fig. 14 g).
The basidiospores may be thin-walled and adapted for im mediate germination, and this is the case of the Protobasidiomy cetes and in some of the lower families of the Autobasidiomycetes. These thin-walled spores when germinated in water or nutrient solutions may sprout like yeast cells and produce gemmae or secondary conidia. In the higher families they are thick-walled and are resting spores. In the majority of Basidiomycetes the spores are discharged from the sterigmata, but only for a short distance (o• 1–o.2 m.m.) . They are, for the most part, distributed by air currents.
Sub Class. Protobasidiomycetes. The Auriculariales is an order of mostly parasitic forms of which the simpler representatives, e.g., Helicobasidium, bear the basidia scattered on a loosely woven mycelium, but in the more typical forms, e.g., Auricularia, the hyphae, which have gelatinous walls, are compacted to form fruit bodies, often of considerable size. The basidia are borne on the upper surface of these fruit bodies and are the terminal portions of hyphae which do not reach the surface, become somewhat wider and divide transversely into four cells. From each of these arises a long sterigma which projects above the surface of the thallus and develops a spore at its tip (fig. 14 e) . The spores can germinate at once and bud off small curved conidia. The fruit body is formed throughout of binucleate cells; the origin of this condition is unknown. The Jew's ear (Auricularia auricula-Judae) is a well-known representative of this order.
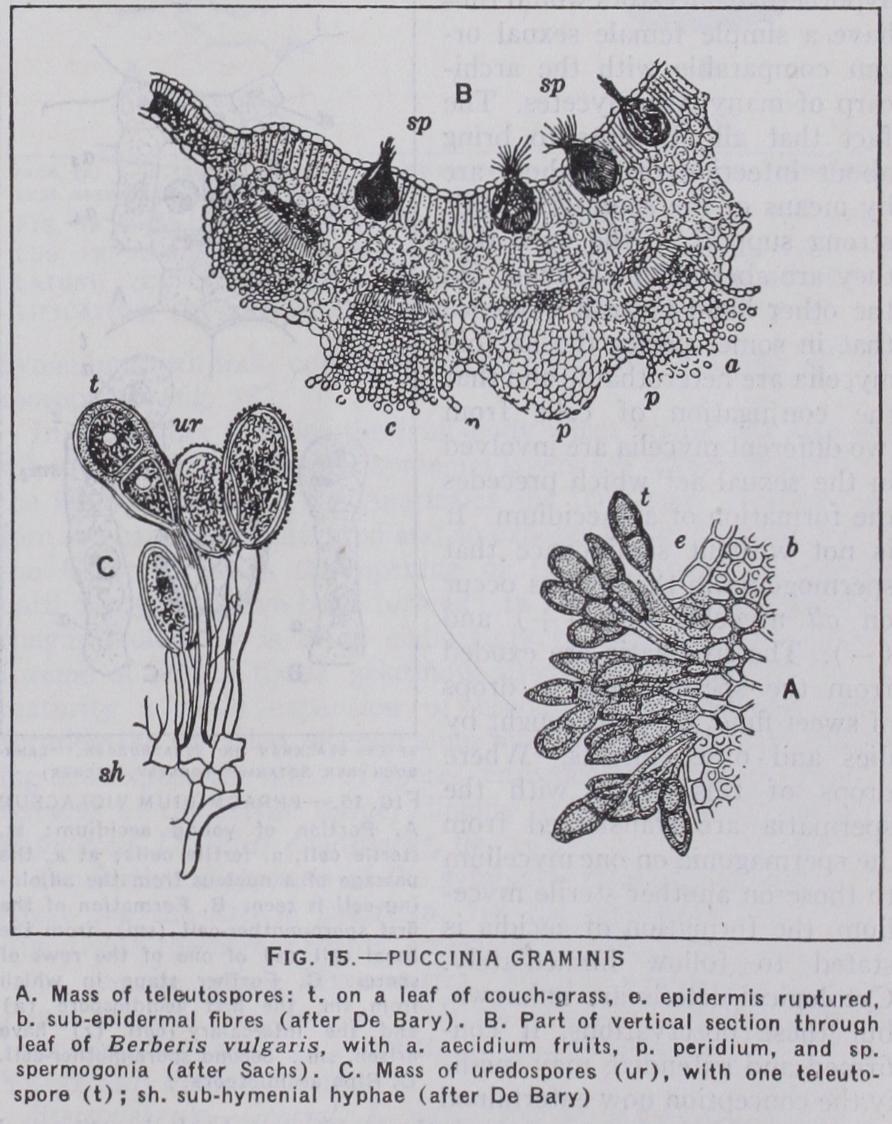
The Uredinales is a large order of parasitic fungi, known as the rusts, which are responsible for the diseases of many of the higher plants—ferns, conifers and flowering plants. Both kinds of mycelium, haploid and diploid are parasitic and may be found on the same host (autoecious) or on different hosts (heteroecious), sometimes belonging to widely different families. They are inter cellular, highly specialised parasites, producing haustoria which penetrate the living cells of the host, and often stimulate the tis sues to increased growth and the formation of galls. They are often short-lived—attacking the leaves of the host and falling with them, but when they extend to 'the stems of perennial plants they are also perennial. No less than five kinds of spore may be produced in the life of one species, viz., aecidiospores, spermatia, uredospores, teleutospores and basidiospores (fig. 15) . The aecidiospores and spermatia are borne on the haploid mycelium and appear early in the year. The first named are borne in fructifi cations known as aecidia, which are cup-shaped receptacles sur rounded and enclosed by an investment (peridium, pseudoper idium), and containing rows of spores, which arise from a layer of cells at the base of the aecidium. The spores are binucleate and it has been shown that in the development of the aecidium, the basal cells are formed from uninucleate cells which, either become binucleate by the migration of a nucleus from a neighbouring cell, or, by uniting in pairs, give rise to a fusion cell with two nuclei (fig. i6). In either case we have a reduced sexual process by which the diploid generation is initiated, the first products of which are the aecidospores. The spermatia are thin-walled cells of small size containing little protoplasm and a relatively large nucleus. They are borne in flask-shaped receptacles, the spermo gonia. As the name indicates, they have been regarded as aber rant male sexual cells. This implies the previous existence of female sexual organs provided with some apparatus for the re ception of spermatia. It has been shown that in certain forms, e.g., Phragmidium violaceum, the fertile cells when young cut off a terminal sterile cell (fig. i 6) which may well represent the re mains of a trichogyne, which in ancestral forms projected above the surface of the host. These hypothetical ancestors would thus have a simple female sexual or gan comparable with the archi carp of many Ascomycetes. The fact that all attempts to bring about infection of the host are by means of the spermatia lends strong support to the view that they are aberrant male cells. On the other hand there is evidence that in some species the haploid mycelia are heterothallic and that the conjugation of cells from two different mycelia are involved in the sexual act which precedes the formation of an aecidium. It is not without significance that spermogonia in these cases occur on all mycelia both (+) and ( — ) . The spermatia are exuded from the spermagonia in drops of sweet fluid, which is sought by flies and other insects. Where drops of this liquid with the spermatia are transferred from the spermagonia on one mycelium to those on another sterile myce lium, the formation of aecidia is stated to follow immediately. Cytological details are unknown, but these observations, if con firmed and extended, may modi fy the conception now entertained of the morphological nature of the spermatia both in this and other groups in which they occur.
The aecidiospores germinate readily on the surface of the appropriate host and the germ tubes enter the tissues by way of the stomata. They give rise to a binucleate mycelium and on this in due course appear the uredospores. These are so-called "sum mer" spores, i.e., conidia of the diploid mycelium, and can infect fresh hosts and so spread the disease. Later in the year and on the same mycelium teleutospores are formed. These are typically thick-walled and resting spores. The uredospores are binucleate and so are the young teleutospores, but in the latter the two nuclei fuse before their development is completed. The teleuto spores are thus the probasidia. They germinate in due course and give rise to the promycelium—the basidium proper, on which after its division into four cells the f our basidiospores, known as sporidia—are produced. These are uninucleate and can at once infect the proper host and develop the haploid mycelium. This is the life history of one of the most complicated forms, the so called eu- forms of which the black rust of wheat (Puccinia gram inis) is an example. In this species the host of the haploid my celium is the common barberry (fig. 15). In other species one or more of the spore types may be absent. The -opsis forms have no uredospores (Gymnosporangiurn sp.). In the brachy- forms the aecidia are absent but the spermogonia are accompanied by sori of uredospores, which in some cases arise as the result of a re duced fertilization, like that already described as taking place at the base of the aecidium. These primary uredospores thus appear to be homologous with aecidiospores. Other species have neither aecidia (or primary uredospores) nor spermogonia and are known as hemi- forms ; there are micro- forms in which teleutospores alone occur. In the latter the binucleate condition is estab lished at an early stage—probably by the migration of a nucleus from one cell to another of the haploid mycelium. The most reasonable interpretation of these forms with a shortened life history is that they have been derived from the more complete forms. The teleutospore is the spore form almost invariably pres ent and the classification of the family is based largely on its structural peculiarities.
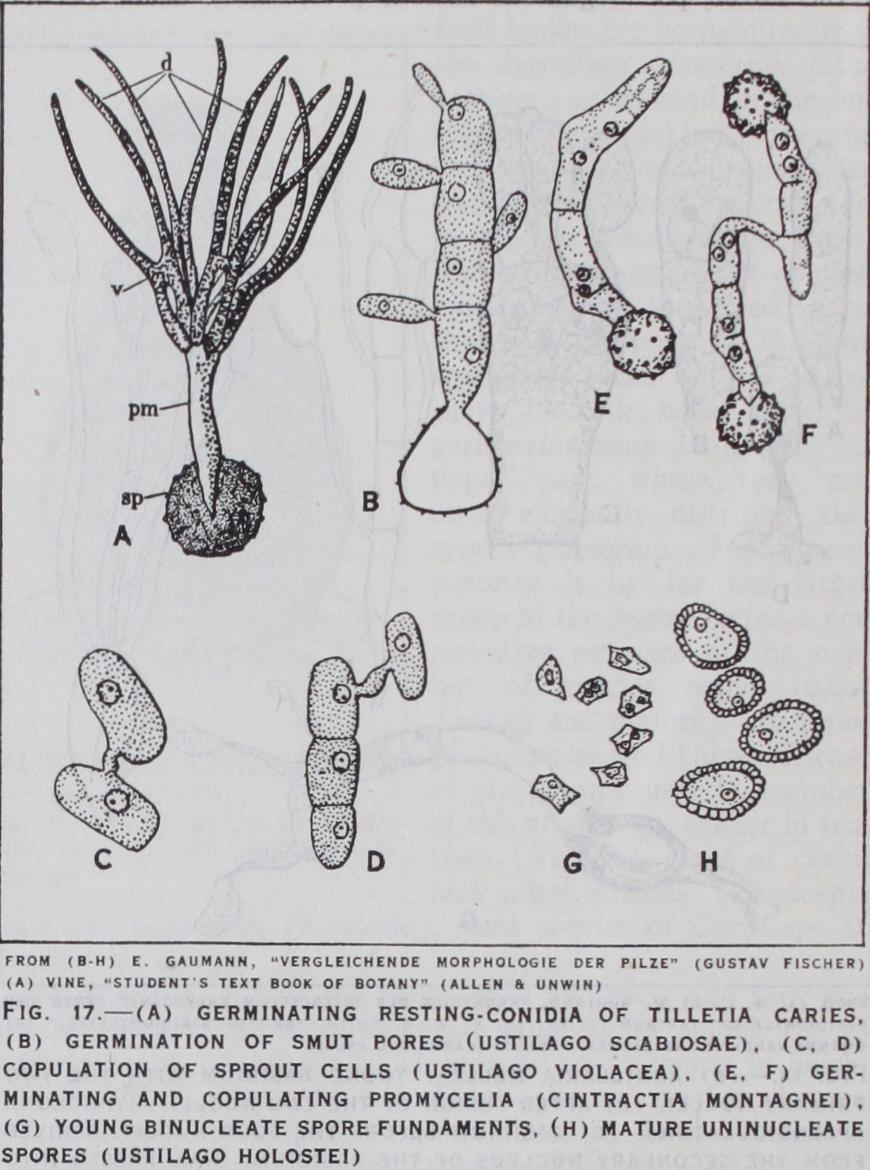
The Ustilaginales were regarded by Brefeld as showing a half way stage in the evolution of the Basidiomycetes from the lower fungi and he placed them in a separate group, the Hemibasidio mycetes. The cytology of the group, however, leaves little doubt that they are true Basidiomycetes and form a parallel group to the Uredinales.
They are parasites on grain and fodder plants causing the well-known diseases smut and bunt. The mycelium, which is usually binucleate, often develops slowly at first and then grows rapidly at a particular stage of development of the host and pro duces its spores, e.g., the smut of wheat (Ustilago tritici) attacks the wheat in the flower. The grains already infected are sown, the plant develops without external sign of disease and not until the ears are formed does the fungus develop in the flowers, and gives rise to a black powdery mass of spores. These chlamydospores are formed from binucleate hyphae which divide into short cells which separate, round themselves off and form the thick-walled chlamydospores. The two nuclei fuse in the spore which is, like the teleutospore of the host, uninucleate (fig. 17).
On germination the spores give rise to a promycelium. In the Ustilaginaceae the nucleus undergoes two successive divisions as in the Uredinales, and the promycelium (basidium) is divided by transverse walls into four cells. These may not form sporidia but may conjugate in pairs by clamp connections or short con jugating tubes (Ustilago nuda) (fig. 17), or they bud off small sporidia, which sprout like yeast cells and eventually conjugate in pairs (fig. i7). The binucleate cells, however formed, can sprout secondary cells or give rise to hyphae, either of which can infect a new host. In one species, Ustilago Zeae, the mycelium in the host is uninucleate and conjugation takes place at the appropriate time between cells of the hyphae, which then round off and form spores.
In the Tilletiaceae, the promycelium remains undivided, eight or more daughter nuclei are formed by division and a whorl of long curved sporidia arise at the tip of the promycelium. These fuse together in pairs, and from the fusion cell secondary sporidia may arise (fig. 17 a).
The Tremellales are a small family of saprophytic fungi with gelatinous fruit-bodies. Their mycelium is binucleate and gives rise to nearly spherical basidia. In these the fusion nucleus under goes the usual divisions and the cell becomes divided by four vertical walls into four daughters from each of which a long sterigma grows up to the surface of the fruit body and develops a spore (fig. 14 f).
The general characters of the sub-class Autobasidiomycetes have already been outlined. Their basidia are undivided and they are typically collected together in a continuous layer, the hy menium, in which are associated with them sterile hyphae or para physes. In the lower families the hymenium is borne on the sur face of the fruit body. This may be a flat crust of indeterminate form and extent or, in species growing on vertical trunks of trees, may be bracket-shaped. In this case the hymenium is on the lower surface. A whole series of forms show that the bracket may narrow at its point of attachment and form a stalk and the fruit body is then differentiated into the stalk or stipe and the broad cap or pileus. More advanced forms show the central stalk and cap-like pileus characteristic of the common mushroom and its relations the toad-stools. A further modification of the hymenium-bearing surface is the development of spines and wrinkles, folds, thin plates (gills) and of pits and tubes; all of which have the effect of increasing manyfold the surface of the hymenium, without correspondingly increasing the size of the sporophore (fig. 18) .
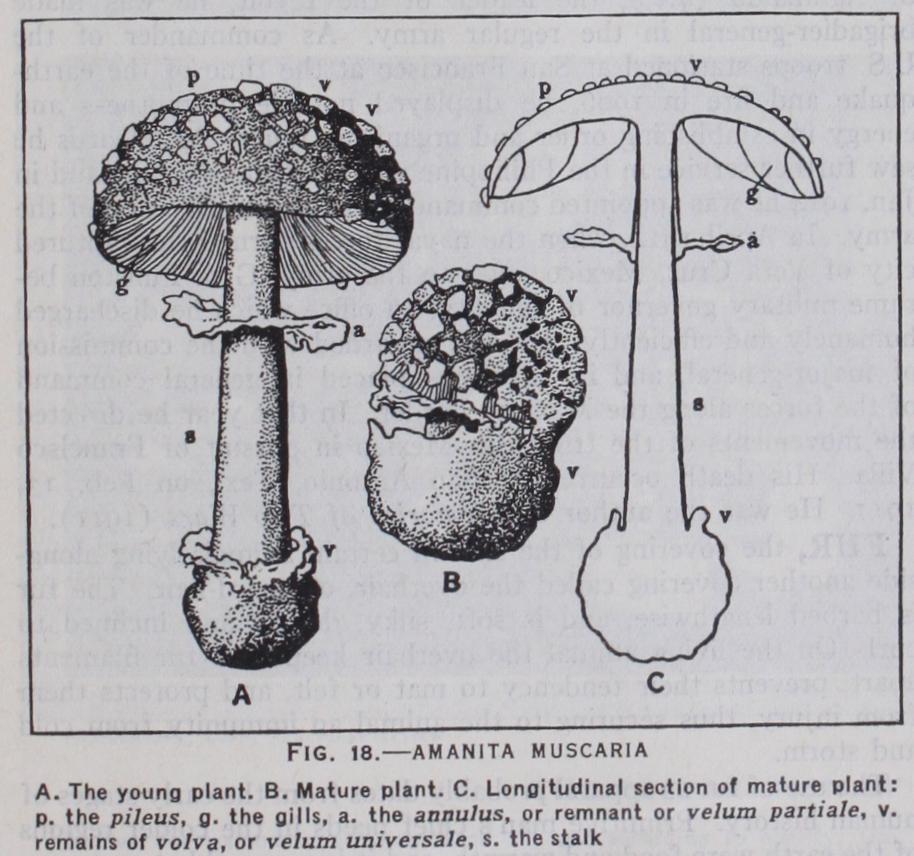
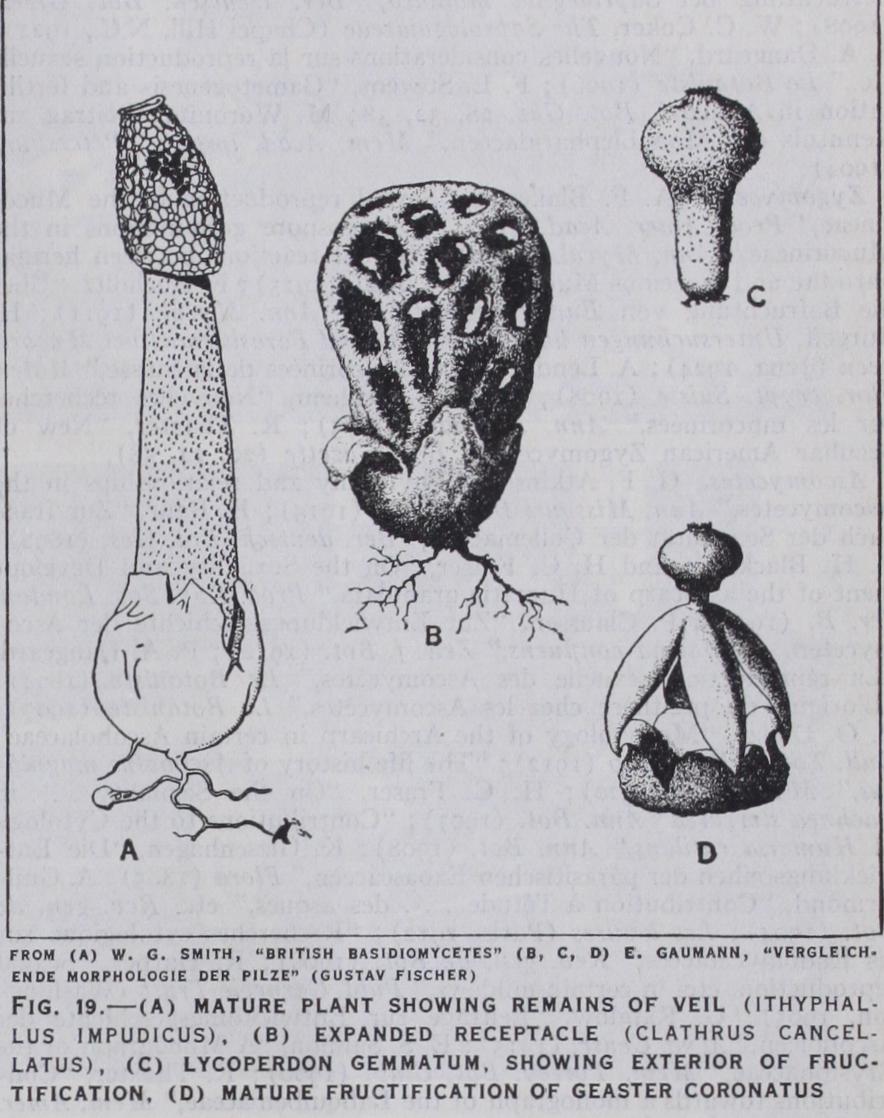
In the higher families the fruit-body when young is a rounded spherical structure and the hymenial layers are differentiated from the inner tissues. In the Agaricales the fruit-body ruptures be fore the basidia are matured and so exposes the hymenium, but in the Gasteromycetes the opening of the fruit-body is delayed until the spores have been formed. In this last-named group the fungus fruit-body is often elaborately constructed and may be formed of fibrous tissue, gelatinous tissue, plectenchyma, etc. At maturity a rapid expansion of pseudo-parenchymatous tissue forming the so-called receptaculum may rupture the outer cover ing and expose the mass of spores, embedded in the mucilaginous remains of the fertile tissue, clinging to its surface (Itliyphallus impudicus, Clathrus cancellatus, fig. 19).
For further discussion of the symbiotic relations of Fungi with Algae see the article LICHENS ; for the symbiotic, relations of Fungi with the higher plants, especially orchids, heaths and forest trees, see the article MYCORRHIZA. For the role of the parasitic Fungi in plant diseases see PLANTS, Plant Pathology. See also