Maturation of Parthenogenetic Eggs
polar and eighty-four
MATURATION OF PARTHENOGENETIC EGGS The maturation of eggs that develop without fertilization is a subject of special interest, partly because of its bearing on the general theory of fertilization, partly because it is here, as I believe, that one of the strongest supports is found for the hypothesis of the individuality of chromosomes. In an early article by Minot ('77) on the theoretical meaning of maturation the suggestion is made that parthenogenesis may be due to failure on the part of the egg to form the polar bodies, the egg-nucleus thus remaining hermaphrodite, and hence capable of development without fertilization. This suggestion forms the germ of all later theories of parthenogenesis. Balfour ('80) suggested that the function of forming polar cells has been acquired by the ovum for the express purpose of preventing parthenogenesis, and a nearly similar view was afterwards maintained by Van These authors assumed accordingly that in parthenogenetic eggs no polar bodies are formed. Weismann ('86) soon discovered, however, that the parthenogenetic eggs of Polyphemus (one of the Daphnidae) produce a single polar-body. This observation was quickly followed by the still more significant discovery by Blochmann ('88) that in Aphis the parthenogenetic eggs produce a single polar body while the fertilized eggs produce two. Weismann was able to determine the same fact in ostracodes and rotifers, and was thus led to the views which later researches have entirely confirmed, that it is the second polar body that is of special significance in parthenogenesis. Blochmann observed that in insects the polar bodies were not actually thrown out of the egg, but remained embedded in its substance near the periphery. At the same time Boveri ('87, x) discovered that in Ascaris the second polar body might in exceptional cases remain in the egg and there give rise to a resting-nucleus indistinguishable from the egg-nucleus or sperm-nucleus. He was thus led to the interesting suggestion that parthenogenesis might be due to the retention of the second polar body in the egg and its union with the egg-nucleus. " The second polar body would thus, in a certain sense, assume the role of the spermatozoon, and it might not without reason be said : Parthenogenesis is the result of fertilization by the second polar body." 8
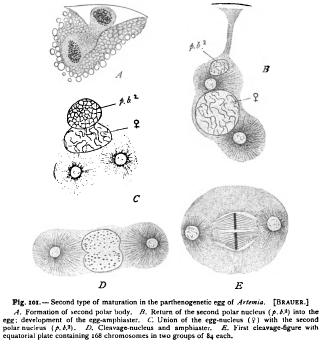
This conclusion received a brilliant confirmation through the observations of Brauer ('93) on the parthenogenetic egg of Artemia, though it appeared that Boveri arrived at only a part of the truth. Blochmann ('88-89) had found that in the parthenogenetic eggs of the honey-bee, two polar-bodies are formed, and Platner discovered the same fact in the butterfly Liparis (89) —a fact which seemed to contradict Boveri's hypothesis. Brauer's beautiful researches resolved the contradiction by showing that there are two types of parthenogenesis which may occur in the same animal. In the one case Boveri's conception is exactly realized, while the other is easily brought into relation with it.

(a) In both modes typical tetrads are formed in the germ-nucleus to the number of eighty-four. In the first and more frequent case (Fig. too) but one polar body is formed, which removes eighty-four dyads, leaving eighty-four in the egg. There may be an abortive attempt to form a second polar spindle, but no division results, and the eighty-four dyads give rise to a reticular cleavage-nucleus. From this arise eighty-four thread-like chromosomes, and the same number appears in later (b) It is the second and rarer mode that realizes Boveri's conception (Fig. 1m). Both polar bodies are formed, the first removing eighty-four dyads and leaving the same number in the egg. In the formation of the second, the eighty-four dyads are halved to form two daughter-groups, each containing eighty-four single chromosomes. Roth these groups remain in the egg, and each gives rise to a single reticular nucleus, as described by Boveri in Ascaris. These two nuclei place themselves side by side in the cleavage figure, and give rise each to eighty-four chromosomes, precisely like two germ-nuclei in ordinary fertilization. The one hundred and sixty-eight chromosomes split lengthwise, and are distributed in the usual manner, and reappear in the same number in all later stages. In other words, the second polar body here plays the part of a sperm-nucleus, precisely as maintained by Boveri.