
ZOA and SCYPHOZOA.
2. The Medusa. Those Coelen terates popularly known as "jelly fish" are scientifically christened medusae, the name referring to the resemblance borne by the long mobile tentacles of some of these animals to the writhing, snaky tresses of the Gorgon Me dusa. The body of a medusa (fig. 3) is shaped like an um brella, and hanging down inside it from the point at which the handle of the umbrella would be attached, is a structure known as the manubrium, at the end of which is the mouth. In this case the tentacles form a circlet round the margin of the umbrella. For details of the organization of Coelenterate medusae reference should be made to the articles HYDROZOA and SCYPHOZOA.
Polymorphism and the Formation of Colonies.—Another marked characteristic of the Coelenterata is their tendency to form colonies of individuals united to each other by a common stem, plate or mass of intermediate tissue. The colonies so formed are generally attached permanently to foreign surfaces, but in some cases they form strings or aggregations of individuals which float or are propelled through the sea. Coelenterate colonies are fre quently characterized moreover by the production in one and the same colony of different kinds of individuals, among which the various functions are distributed. There may co-exist in a single colony not only polyps as well as medusae (fig. 4), but more than one kind of each of these types of individuals. There may be sex ual medusae and medusae trans formed into locomotory organs; polyps whose main f unction is digestion and others which catch and paralyse prey; and so forth. The phenomenon thus outlined is known as polymorphism and metagenesis is a form of it ; it is one of the most interesting mani festations of the diversity of ani mal life, and is considered in more detail at the end of the article HYDROZOA.
5). In this species the eggs are shed into the sea by the parent and are there fertilized. After fertilization the egg divides into halves and each of these once more into two. The four cells (or blastomeres) so formed are equal or sub-equal in size. This process of subdivision is continued until a considerable number of small cells have been formed, and these are so arranged as to constitute a hollow sphere containing fluid. The sphere is known as a blastula and the cavity as a blastocoele. The blastula con tinues to develop in such a way that one side of the sphere first flattens and then becomes concave, being finally completely in vaginated into the other half. This process practically obliterates the blastocoele, and forms in its stead a second internal cavity, the archenteron, which communicates with the exterior by means of a small opening, the blastopore. There has thus been produced a typical gastrula such as has been previously described ; its archen teron becomes directly transformed into the coelenteron of the adult, and the blastopore becomes the adult mouth. During its early stages the animal moves through the water by the aid of cilia which appear on its outer surface during the blastula stage, but before it begins to assume the adult shape it tends to remain at the bottom, moving horizontally and resting from time to time and often becoming temporarily attached to the bottom mouth downwards, during which time it may execute slow creeping movements. From this larva the adult anemone-polyp is achieved by degrees. At a given time the larva settles down and attaches itself to a foreign surface ; its proportions change, it acquires ten tacles, and becomes a small polyp.

The embryology of the Coelenterata varies considerably from one species to another, but generally speaking it is characterized by the presence of a gastrula-stage or of some equivalent. Fre quently no actual gastrula exists but instead a corresponding stage known as planula is produced, in which there is no blasto pore, the embryo consisting either of a hollow two-layered organ ism or of a central mass of endoderm-cells covered externally by a layer of ectoderm (fig. 5) . Planulae and gastrulae may be free-swimming organisms moving by means of cilia, or may be contained within the coelenteron of the parent or within a pro tective enclosure so that they have no actual free existence, and are born as young polyps.
Motion.—The movements of which a Coelenterate is capable depend upon its shape, organization and mode of life. A jellyfish is an active free-swimming creature which progresses through the water as a result of rhythmic or spasmodic contractions of its bell-like body brought about by the action of muscles. Different kinds of jellyfish are active in varying degrees; some are much stronger swimmers than others, some are decidedly sluggish, whilst a few are creepers. Movement in polyps varies according to whether they possess a skeleton or not ; in those provided with such a structure movement is naturally limited to those actions which the tentacles or other parts of the polyp can perform unre stricted by the hard parts. If a polyp is free from skeleton, it can move in a variety of ways; rarely, it swims actively by concerted lashing movements of the tentacles ; sometimes it will creep on these organs ; or it may attach itself by the base, bend over and attach the tentacles, then loosen the base, move its body and re-attach the base elsewhere. On the other hand it may simply loosen its hold and allow the motion of the water to carry it else where, and in such case may inflate itself with water in order to become more buoyant; or it may creep upon its base, a method characteristic of many of the large polyps of the sea anemones.
The movements exhibited, apart from those connected with actual locomotion, consist of contractions and expansions of the tentacles and body, movements of the mouth, and so forth, con nected with capture and swallowing of food, with retraction for the purpose of sheltering from adverse conditions, and with simi lar matters. Tentacles are usually highly contractile, and in their most concentrated condition are very much smaller than when expanded. The whole body may be relatively rigid and not, as a whole, very contractile, in forms possessing a high development of the mesogloea ; but in many cases it is as contractile as the tentacles and may be reduced at need to a bulk very much smaller than its size when expanded; e.g., to an eighth of its maximum bulk.
Muscles.—Although the general substance of a Coelenterate is contractile, the definite movements which it performs are the result of muscular or ciliary action. The muscular system in its simplest condition consists of a single layer or sheet of muscle fibres, lying on the inner or outer surface of the mesogloea. The muscle-sheets of Coelenterates are normally only one fibre thick, and each fibre lies directly upon a supporting surface of meso gloea. Consequently if strong localized muscles (as distinct from diffuse sheets) are required, these are generally attained by the simple expedient of pleating the surface of the mesogloea into ridges, on the surface of which lie the fibres, still in a single layer. In certain cases muscle becomes embedded in the mesogloea. The individual fibres are not in the simpler instances independent structures, but belong to and form part of epithelial cells of the ectoderm or endoderm. Thus the combination of each cell with its "muscle tail" (or tails) constitutes a musculo-epithelial cell, a structure characteristic of Coelenterata but rare among higher animals (fig. 8). In more specialized cases the muscle-fibres of the ectoderm become separate structures, the epithelial cells which produced them being of insignificant proportions.
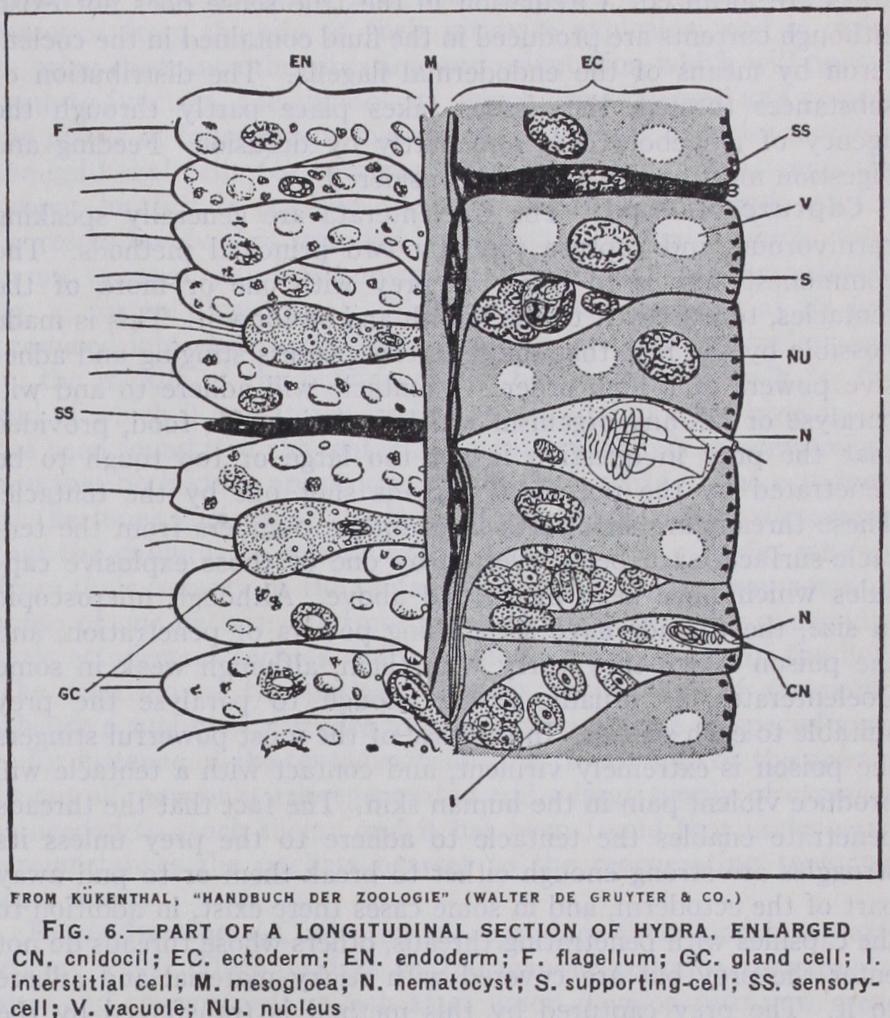
The Finer Structure of the Tissues.—The structure of the layers of the body must now receive short notice (fig. 6). The mesogloea is a sheet of very variable structure ; sometimes it is gelatinous in texture, sometimes almost cartilaginous, or again it may be very watery and unsubstantial. It contains within its substance cells which have wandered there from the other layers, and the functions of which are not fully understood. Some of them at least are amoeboid cells which carry or transmit nutri ment from the endoderm (the digestive layer) to other parts of the body. Others probably add to the mesogloeal substance by secretion, whilst it is possible that some of them constitute part of a mesogloeal nervous system. At its highest development the mesogloea possesses fibres as well as cells, but these are not at all of the same nature as the muscular fibres. The cell-layers are best considered separately, and the following details apply to a sea anemone, which will serve as a suitable example. The ecto derm is an epithelium consisting of elongated supporting cells which are ciliated in some parts of the body, and among these occur large numbers of cells of other kinds. There are glandular cells which secrete mucus, etc., sensory cells which constitute the peripheral receptive elements of the nervous system, cnido blasts which secrete curious explosive capsules about which more will be related shortly, and undifferentiated interstitial cells which may give rise to other kinds at need. In the deeper parts of the epithelium lie nerve cells (the nervous system is further described below) and on the surface of the mesogloea, in given parts of the body, are the muscle fibres, which are here inde pendent of the supporting cells. The endoderm resembles the ectoderm in general constitution, but its supporting cells, instead of being ciliated, bear each a single large cilium or flagellum, and each possesses a basal muscle fibre, and so constitutes an epithelio muscular cell. Many of them are digestive in function. Gland cells are to be found in the endoderm, as also are sensory, nervous and interstitial cells ; cnidoblasts are most characteristic of the ectoderm, but also occur freely in the endoderm where necessary.
The germ cells of the Coelenterata, those cells which will in due course form eggs or spermatozoa, are formed sometimes from interstitial cells, sometimes from differentiated epithelial cells. They may originate in either ectoderm or endoderm, but are not necessarily first formed in the region in which they will ripen.
There is another method of feeding which may supplement or replace the characteristic one. Small food-particles which come in contact with some part of a Coelenterate become entangled in slime secreted by that part, and these are transported by means of cilia acting in definite directions to the mouth. This method of feeding is probably the predominating one—for instance, in certain sea anemones—and a good example of its working may be seen in the jellyfish Aurelia. (See article SCYPHOZOA for an illustration.) When Aurelia is feeding, planktonic organisms be come entangled in strings of slime on the outside of the swim ming-bell, and are conveyed by the movements of the bell aided by the action of cilia to the edge of the umbrella. Arrived at this point they are concentrated into eight masses at points situated at regular intervals round the margin of the bell. From time to time these masses are licked off by one or other of the four large arm-like processes which hang down under the bell round the mouth; and since each of these arms contains a groove leading directly into the mouth, the food-mass is from this point onwards easily transported into the stomach. The underside of the swim ming-bell also contributes to the eight food-masses and the oral arms themselves collect plankton independently. This seems to be a normal mode of feeding in Aurelia, but at the same time it can and does, at any rate when young, catch fishes and other organisms as well as plankton.
The structure of both cnidae and cnidoblasts varies greatly from one Coelenterate to another, and both may be complex. The cnidoblast possesses, in a number of cases at least, contractile fibres of varying disposition, and may also contain a coiled spring like structure (the lasso) which probably resists the tearing away of the nematoblast from the tissues by a struggling prey. In a typical cnida the thread is provided with a series of spirally arranged barbs which help to fix it in the tissues of the prey, and bears with it poison which enters by the wound it has made. This kind of cnida, which is very frequent, is known as a nematocyst, in contrast to another type common in sea anemones and known as a spirocyst. The latter has no armature, but the outer surface of its evaginated thread has strongly adhesive properties, and is probably not poisonous.
True cnidae occur nowhere in the animal kingdom save in the Coelenterata. There are similar structures developed in at least two other kinds of animals (the Myxosporidean Protozoa and certain Nemertine worms), but they are not exactly the same; a number of other structures which have been confused with nema tocysts are not actually such. The most interesting cases which have been investigated in this connection are those of certain animals in whose tissues actual Coelenterate nematocysts are pres ent in an unexploded and functional condition. It has been con clusively shown that an animal may feed upon a Coelenterate, thus swallowing quantities of nematocysts, and that a number of these may pass unexploded through its food-canal, and may sub sequently become arranged in a definite manner in its tissues. The best known instance of this is provided by certain of the Nudibranch molluscs or sea slugs. In some cases at least these stolen nematocysts appear to be of value to their host.
Nematocysts and spirocysts, which are produced by interstitial cells, are not developed in the place where they will function. In some cases this means simply that they develop in the deeper layers of the epithelium and move up to the surface as they mature. In other cases however a much more interesting story is involved. The capsules are here secreted by cnidoblasts far removed from the site of their ultimate explosion, and in order to bring them into the situation and orientation which will permit them to function, the cnidoblasts, which are amoeboid and possess the power of independent movement, drag their capsules between the epithelial cells (or transport them by other means) until the proper locality is reached. A most interesting example of the processes involved is supplied by the state of affairs which exists in an unusual jellyfish known as Haliclystus (see SCYPHOZOA, fig. 4, for an illustration). This animal has the edge of its bell produced into eight arms, each tipped by a tuft of tentacles; and on the middle of the outside of the bell it bears a stalk by the end of which it can attach itself to seaweed and other objects. It has been found that the tentacles contain two kinds of nematocyst, and that both kinds are produced in definite parts of the ectoderm of the inner surface of the bell. It has further been discovered that the cnidoblasts containing the two kinds of capsules migrate from their nursery to the tentacles; but that on the way some of those of one pattern (and not those of the other) disappear into little ectodermal pockets which lie close to the margin of the bell. The function of the pockets appears to be that of reservoirs whence a supply of capsules may be drawn in case of special need —for instance, if the tentacles belonging to one side of the animal be cut off, new ones are regenerated and a large supply of capsules is needed to stock them ; and it has been found that under such circumstances the pockets nearest to the regenerating tentacles were empty.
The following experiment is a case in point. Three shore-crabs (Carcinus maenas), three common hermit-crabs (Eupagurus bernhardus) and three of the hermits proper to Adamsia (E. prideauxi) were selected, and into the body-cavity of each was injected o•Ic.c. of a filtered maceration of Adamsia. At the end of three minutes the bernhardi were completely paralysed after a short phase of tetanization; and at the end of an hour they were dead. The shore-crabs died between five and nine hours after having exhibited the symptoms characteristic of this poison. On the other hand the prideauxi showed no apparent trouble ; they retained all their agility and all the accustomed vivacity of their movements. At the end of 24 hours their condition was normal and they survived indefinitely. Other experiments confirm the result of this one, and the interesting fact emerges that E. pri deauxi is immune from the poison of its anemone unless injected in inordinately large amount. Thus the association between the two animals includes a physiological as well as a morphological adaptation. It has been shown further that the serum of E. prideauxi is able to neutralize the Adamsia poison and that if a shore-crab be inoculated with prideauxi serum, it can afterwards withstand and recover from a dose of poison which would other wise be fatal. It is probable that E. prideauxi gains its immunity in the first instance as a result of the fact that its close association with the anemone involves the constant entanglement of small portions of the latter, or of stinging cells derived from it, in the hermit's food. The intestinal content of the hermit usually con tains such fragments of the anemone. Although in these experi ments the poison of Adamsia did no harm to other anemones, yet in other cases one anemone may sting another, even of the same kind, and produce a wound which may prove fatal—so that an anemone is not necessarily immune from poison produced by its own kind. It is possible, however, that the injured animal in such cases is, to begin with, in a poor condition.
Development: E. W. MacBride, "Invertebrata," in W. Heape, Text-Book of Embryology, vol. i. (1914). Histology: K. C. Schneider, Lehrbuch der V ergleichenden Histologie der Tiere (Jena, 1902) and Histologisches Praktikum (Jena, 1908). Nematocysts: L. Will, Natur forschenden Gesellschaft (Rostock, 1909-1o) ; P. Schulze, Archiv fur Zellforschung, with bibl. (ed. R. Goldschmidt, Leipzig, 1922) ; and in animals other than Coelenterata: C. H. Martin, Biologische Centralblatt (ed. J. Rosenthal, Erlangen, Leipzig, 1914). Nervous System and Behaviour: G. H. Parker, The Elementary Nervous System (1919). Digestion: H. Boschma, Biological Bulletin (Woods Hole, Mass., 1925) ; R. Beutler, Zeiischr. f iir vergl. Physiol. (1924-26) . See also H. Boschma, etc., for Zooxanthellae. For recent lists of literature see W. Kiikenthal's Handbuch (supra), and for further references the articles HYDROZOA, ANTHOZOA and GRAPTOLITES. (T. A. S.)