Lamellibranchia
filaments, ganglia, found, chamber, lamellae, organs, eyes, gills and separate
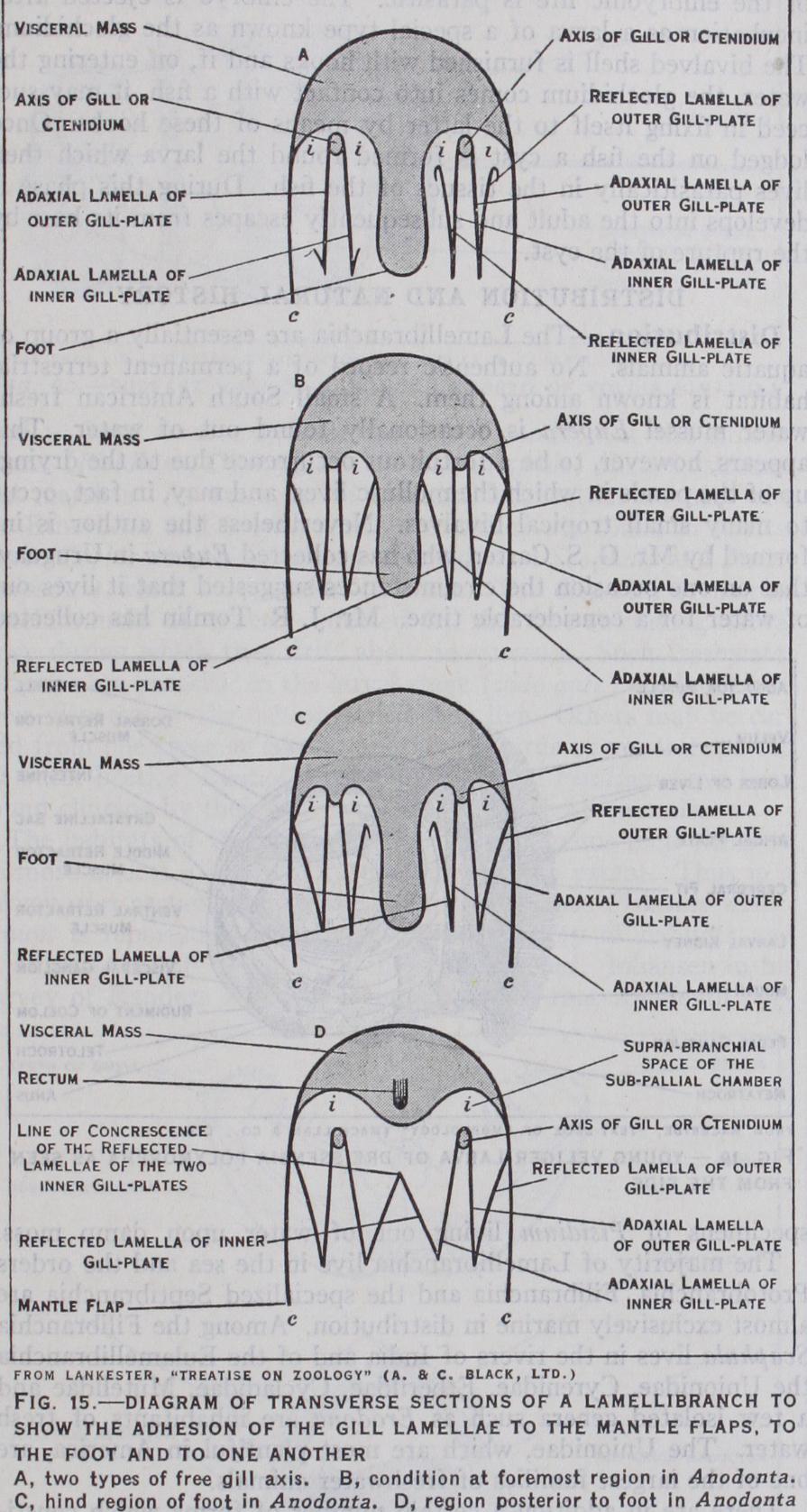
I. The rows of filaments, instead of being opposed as they are in the Protobranchia, become parallel to each other and hang downwards in the mantle-cavity and the extremities of the individ ual filaments are bent upwards so that each row of filaments is doubled. The gill thus consists of four series of filaments (la mellae). Of these the two lamellae adjacent to the axis are called the direct lamellae, those formed by the upturned portions of the filaments being known as the reflected lamellae.
2. The next advance in complexity is the development of discs of cilia which face each other on adjacent filaments. The masses of cilia of the opposed discs fit into each other rather like the bristles of two brushes, and the discs thus lock the adjacent fila ments together.
3. The direct and reflected lamellae are joined by "interlamellar junctions." This stage of development is found in the Filibranchia (e.g., Pecten).
In the Eulamellibranchia there are vascular interlamellar and interfilamentary junc tions.
4. The lamellae may be thrown into folds and the extremities of the filaments which are free in primitive forms may fuse with the mantle. The result is that the mantle-cavity becomes subdivided by the partition thus formed by the gills into an upper and a lower chamber and the respiratory and nutritive currents of water are canalized. The water enters at the inhalant orifice, passes into the lower chamber, thence through the meshwork of filaments into the upper chamber, and is expelled by the exhalant siphon.
5. In the Septibranchia the gills are profoundly modified. Their respiratory function is suppressed by the development of muscular tissue in their substance and the gills are converted into a con tractile partition.
The role of the gills in providing an incubatory chamber is expanded in another section of this article.
Coelom and Excretory Organs.
In the Protobranchia the primitive connection between the pericardium and the gonadial coelom is retained as the reproductive organs open into the reno pericardial canals. In the Filibranchia, such as Pecten, the genera tive ducts open into the kidneys, and in the Eulamellibranchia they have separate orifices. The kidneys are symmetrical and ramified and in certain specialized forms they communicate with each other. The excretion of hippuric acid is carried out by the pericardial glands.
Nervous System.
The nervous system consists of pairs of ganglia (connected by commissures), nerve cords and sense organs.
With one exception the ganglia are separate from each other and the extreme condensation of the nervous system arising from the juxtaposition of the ganglia which is found in Gastropoda and Cephalopoda is not found. In the Protobranchia the pleural, cere bral, pedal and visceral ganglia are all distinct. In all other La mellibranchia the cerebral and pleural ganglia are fused together.
The visceral commissure is always long. The pedal ganglia are closely approximated. The visceral ganglia are usually placed on the surface of the posterior adductor muscle and in most Eula mellibranchia are more or less attached to each other. There is no stomatogastric system such as is found in the Gastropoda, the alimentary canal being innervated from the visceral commissure. Sense organs consist of eyes, otocysts, osphradia and tactile or gans. In a very few genera eyes are found on the head (e.g., in the Mytilidae). It is to be inferred that, in general, eyes in such a position would be useless, as this region is completely covered over by the shell. Eyes are consequently found on parts which can be extended from the latter, viz., on the syphons and the edge of the mantle. In their most simple form these are simple pigment-spots. But in certain groups they are more com plex and in Pecten and Spondylus they attain the maximum corn plexity, consisting of a cornea, conjunctiva, lens, retina, deep pig ment layer and tapetum. The optic nerve divides into two branches just below the optic cup and one branch passes round the latter to enter the retina. The extremities of the retinal cells (rods) are turned towards the interior of the optic globe (i.e., away from the direction in which the light-rays enter the latter).